Today we've got something really special for you! If you've been following along in the PricePlow Discord, you know we've been running monthly podcasts with Glaxon leaders Joey Savage (chief science officer and formulator) and Michael Bischoff (founder and CEO).
Meanwhile, we regularly run Formulator's Corner segments here on the PricePlow Blog, where we dream up exciting theoretical supplement formulations using novel ingredients from innovators like NNB Nutrition.

In Formulator's Corner #9, we publish Joey Savage's design for a Pre-Workout Support Supplement to take the standard pre-workout way further. Of note, there's a massive 2000 milligram dose of BioNMN!
Today, we merge the two, discussing a pre-workout supplement support formula that Joey made for one of our key followers, Dave Cook.
Formulator’s Corner #09: The Ultimate Pre-Workout Support Formula
In one of our recent public discussions in the Discord, we brought Dave "on stage" after winning a supplement trivia contest, and Joey offered to build a stack to help Dave achieve his goals.
The duo ended up building an additive formula to support your existing pre-workout supplement, code named "Super Dave". It consists of a laundry list of incredibly-dosed ingredients selected to complement and round out the industry's standard pre-workout formulas. Meaning, you won't see anything like caffeine, betaine, beta-alanine, or citrulline inside.
Instead, this is a cocktail of other ingredients, and many of them should be considered for next-generation pre-workout supplements. The doses are all huge (eg. 2 grams of both MitoBurn and BioNMN), so no formulator can put all of this into a finished product and keep it affordable, but the effects were so strong for Dave that there's clearly something big happening here.
Consider this a thought experiment that formulators around the globe can pick from to incorporate into their own products so that we don't get stuck with the "same old pre-workout". Some of these ingredients, like BioNMN, are how we believe you can take an ordinary pre-workout to extraordinary.
Let's get into it, but first, be sure to sign up for our future Formulator's Corner news alerts:
Subscribe to PricePlow's Newsletter and Alerts on These Topics
![]()
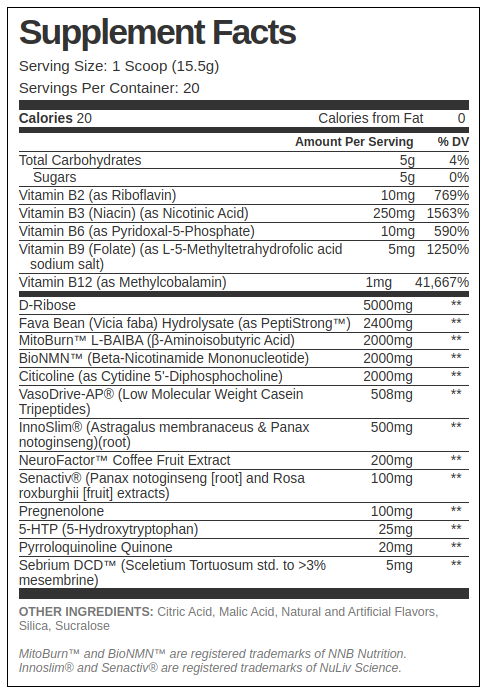
Dave’s Pre-Workout Support Formula Ingredients
In a single 15.5-gram serving of Joey and Dave's Formulator's Corner pre-workout support, you get the following:
-
Ribose – 5,000 milligrams
D-ribose plays an important role in the synthesis of adenosine triphosphate (ATP), your body's fundamental unit of cellular energy. Sometimes ATP is referred to as "molecular currency" because it contains high-energy bonds that can be delivered to provide actual chemical energy for cells.
If your body were an automobile, ATP would be the gasoline – it's that important. Without enough gasoline, a car simply won't run for long. Your body is not so different in that absolutely everything your body does, every single function or task you ask your cells to perform, requires ATP. No ATP means no metabolic function whatsoever.
ATP is made up of three fundamental components: adenosine, phosphate, and ribose.[1] You need all of these to make ATP in adequate quantities.
Ribose is classified as a pentose sugar. The pentose refers to the fact that ribose has five carbon rings. It's produced by a metabolic process called the pentose phosphate pathway (PPP), which is governed by the process of glycolysis.[1]
However, the PPP is sharply rate-limited: it requires access to an enzyme called glucose-6-phosphate dehydrogenase, which tends to be scarce and thus tends to bottleneck the production of ribose. The downstream effect of this can be to stifle ATP production as well.[1]
In fact, research has found that when ribose availability declines, ATP production sharply declines with it.[1] Exercise consumes ATP and hence, consumes ribose, which can then become a logjam on ATP replenishment. Supplementing with ribose has the potential to accelerate ATP replenishment through bypassing the ribose-PPP bottleneck on ATP synthesis.[1]
With some success, ribose has been used to increase physical performance by alleviating pain, managing cramps, and reducing stiffness. It can also help manage the symptoms of persistent conditions like chronic fatigue syndrome (CFS), myocardial dysfunction, and fibromyalgia.[1]
However, this is just the start of our cellular energy play. We have far more coming with MitoBurn, BioNMN, niacin, PQQ, and more. And we still suggest 3-5 grams of creatine each day.
-
Fava Bean (Vicia faba) Hydrolysate (as PeptiStrong) - 2,400 mg
One of the goals for any good pre-workout supplement is to help consumers get the most out of their training, which almost always means maximizing the anabolic response and facilitating muscle growth.
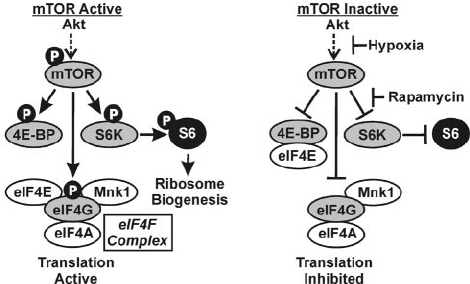
There are a few different strategies for maximizing anabolism. One of the most important is an informational metabolic switch called the mammalian target of rapamycin (mTOR). mTOR is a protein kinase, a type of enzyme that modifies proteins by phosphorylating them.
PeptiStrong is an awesome designer ingredient from Nuritas that helps upregulate mTOR activity.
mTOR in context: what it is and how it works

This is how you shake up a month of training
This topic is complex, to say the least. But in simple terms, mTOR regulates protein synthesis and metabolism in a huge range of organisms. In fact, one of the early mTOR analogs was discovered in yeast, a remarkable finding that shows how the TOR kinase (missing the m here because we're talking about more than just mammals) is strongly evolutionarily conserved.[2]
Whenever a biochemical mechanism appears across such a broad range of species – the basic meaning of evolutionarily conserved – we should take that as a clue that it's really, really important.
The mammalian TOR kinase, mTOR, can cause severe muscle wasting in mammals when its activity is suppressed.[3] Obviously, uncontrolled muscle wasting is a catastrophic outcome for a mammal, one that can easily lead to premature death.
On the flip side, mTOR upregulation can actually prevent muscle loss in mammals, even during involuntarily immobilization, which would ordinarily produce muscle wasting since physical activity is a crucial signal for the body to maintain existing muscle.[4]
Paradoxically, certain mTOR inhibitors like rapamycin can nonetheless extend lifespan and help protect muscle as an organism ages.[5]
The key to mTOR's ultimate effect seems to be dependent on how and for how long it's activated.[5]
There’s more to the story than just mTOR

Dubbed "Super Dave" after the one and only Dave Cook on the PricePlow Discord. Follow Dave on Instagram: @cook.dave10
So how do we resolve the paradox? As it turns out, the specific effect of mTOR activation or inhibition depends on how it works in tandem with other proteins and kinases.
For example, mTOR is but one component of mTOR complex 1 (mTORC1), a cluster of proteins that control muscle growth in response to exercise and/or dietary-protein intake.[6] mTORC1 is the cluster of proteins that actually makes your muscles grow.
Scientists have found that in rodents subjected to long-term resistance training, knocking out mTOR significantly decreases the amount of muscle they build in response to training,[7] and mTOR-knockout rodents lose muscle to wasting very quickly.[8]
So if we want maximum gains, we need to make sure that not just mTOR, but also the entire mTORC1 cluster, is upregulated.
The role of phosphorylation in mTOR activity
When you get down to the biochemical nuts and bolts, you'll see that mTOR activity entails upregulated phosphorylation, the transfer of high-energy phosphate groups between molecules.

The phosphorylation of S6 ribosomal protein is a marker of mTORC1 activity[9] (the P stands for phosphorylation). Phosphorylation of mTOR drives ribosome biogenesis
One corollary of this fact is that in order for mTOR kinase to be maximally effective, we must set it up to be absolutely loaded with phosphate groups. That's why PeptiStrong shows up in Joey's pre-workout support formula. Most pre-workouts have some kind of phosphate donor. Creatine is an excellent example of a phosphate donor.
PeptiStrong upregulates mTORC1 and increases S6 phosphorylation
The way PeptiStrong works is by stimulating mTOR phosphorylation and mTORC1.[10]

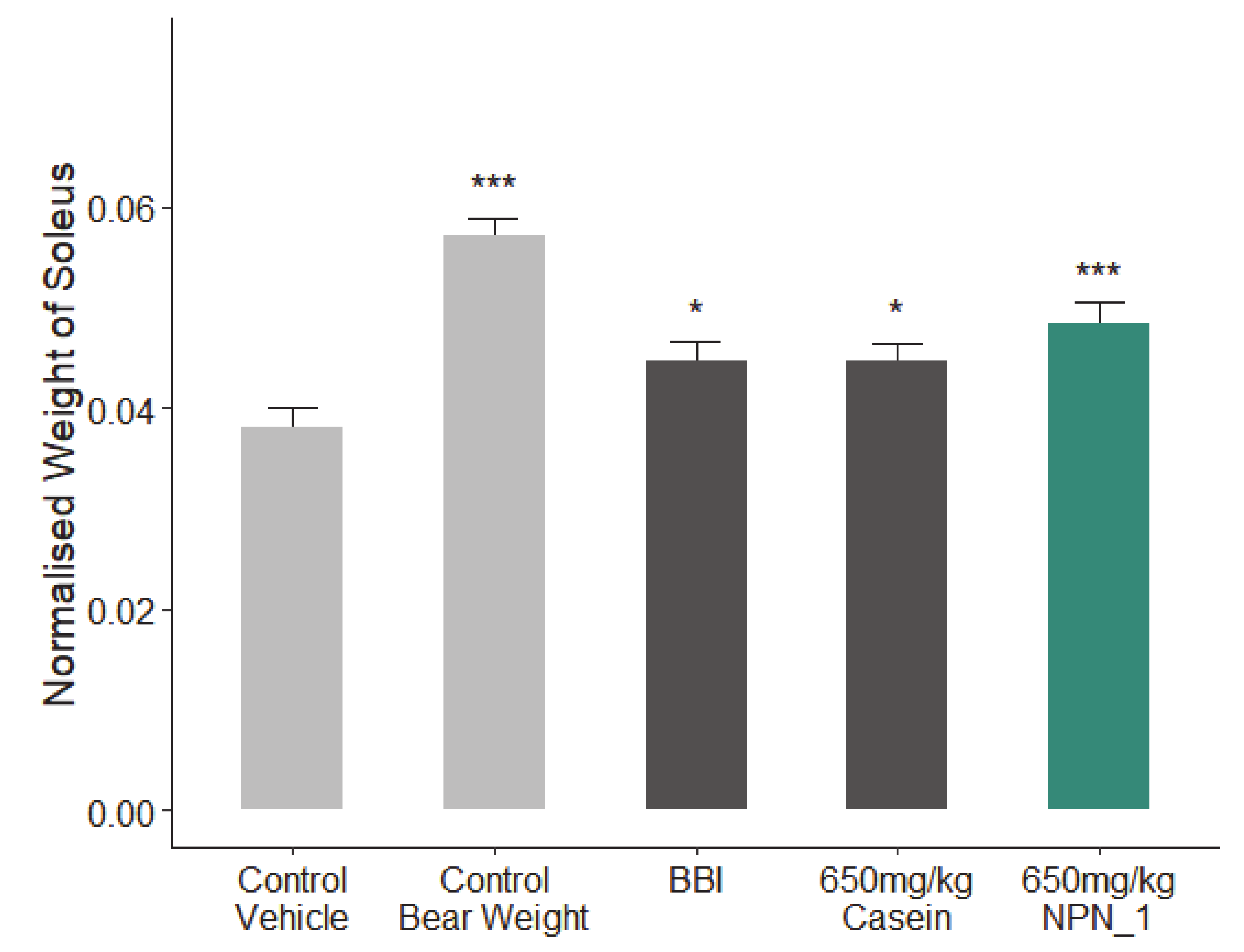
The effect of NPN_1 on muscle mass that would otherwise atrophy[10]
The researchers who have looked at fava bean hydrolysate proteins like PeptiStrong (referred to as Natural Protein Network 1 or NPN_1 in studies) have found that mice treated with NPN_1 have much more S6K1-phosphorylated S6 ribosomal protein[10] – a protein that is expressed by mTOR activity and is a product of mTORC1.
More S6 ribosomal protein = more ribosome synthesis
The reason we love S6 ribosomal protein so much is that it drives a process called ribosome biogenesis, in which your body creates new ribosomes. Just as a reminder, ribosomes are the organelles in your body's cells that are responsible for creating large, complex proteins out of smaller, simpler ones.
In skeletal muscle, the complex proteins created by ribosomes eventually become new muscle tissue, so upregulated ribosomal activity is awesome for muscular hypertrophy.
PeptiStrong reduces inflammation
Ah, inflammation – as most of us have heard, especially over the past decade, it can be a real killer. When it goes uncontrolled, it gives rise to a condition called chronic inflammation, in which it becomes constant and unremitting. Chronic inflammation, in turn, can eventually cause devastating diseases of civilization like type 2 diabetes,[11,12] cardiovascular disease,[13] and others.[13]
Chronic inflammation can also inhibit new muscle growth,[14] by impairing the anabolic response. This can spell disaster for your hard-won gains, but it can also lead to muscle loss,[15,16] leaving you even worse off than before.
The good news is that in animal models, PeptiStrong has been shown to downregulate the inflammatory cytokine tumor necrosis factor alpha (TNF-α).[10] TNF-α tells your body to initiate the inflammatory response, which is why upregulated TNF-α is linked to muscle wasting,[17] including age-related sarcopenia.[18]
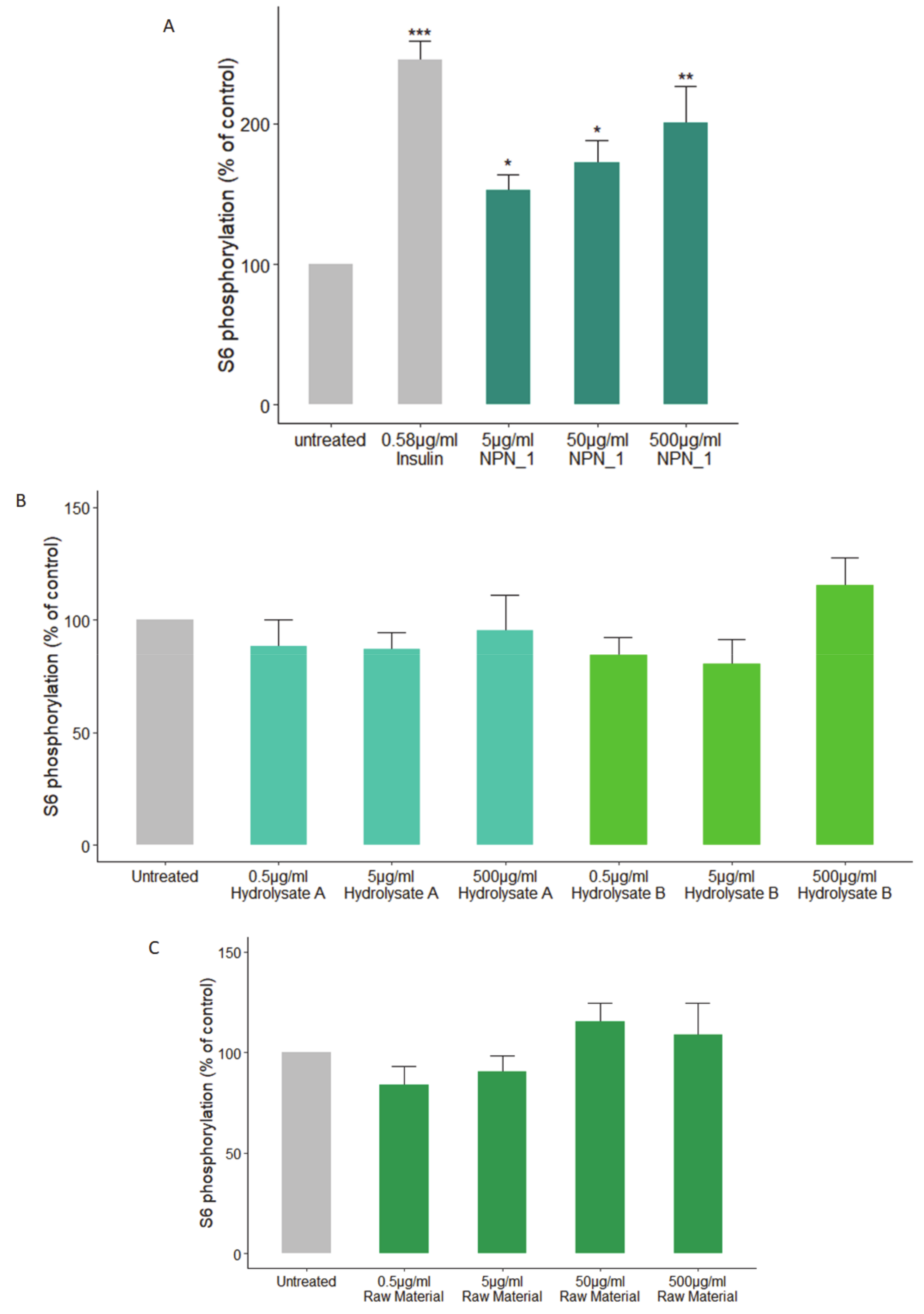
"Effect of the natural peptide network_1 (NPN_1) treatment on S6 phosphorylation. C2C12 cells were treated with (a) NPN_1 (5–500 μg/mL), (b) hydrolysate A, hydrolysate B or (c) raw material (5–500 μg/mL) for 30 min following a starvation protocol and compared to untreated cells (control) and expressed as the % of controls (one-way ANOVA analysis"[10]
PeptiStrong downregulates certain unwanted genes
The research literature on PeptiStrong indicates that it can favorably decrease the expression of two important genes:
- First up is Fbxo32, a gene that must be expressed in order for muscle wasting to take place.[19,20] In Fbxo32-knockout mice, muscular atrophy is markedly reduced.[20]
- The next is Trim63, a gene that drives muscular catabolism during starvation.[21] One thing to note about Trim63 is that it breaks skeletal muscle down in order to supply organs with amino acids during restricted calorie intake.[10] So if you're using PeptiStrong on a cut, be sure to eat plenty of protein.
PeptiStrong vs. traditional protein supplements
In one animal study, researchers gave PeptiStrong or casein to mice with muscular atrophy in their hind legs, and found that the PeptiStrong group actually lost less muscle than the casein group.[10] That's pretty remarkable, considering casein's bona fides as a supplemental protein.
Granted, PeptiStrong is used in far lower doses than casein. Here we have 2.4 grams, whereas it's not uncommon to take 30 grams or more of casein in one sitting.
Still, pound-for-pound, PeptiStrong seems to be more effective, which says something.
-
MitoBurn (L-β-Aminoisobutyric Acid) – 2,000 mg

NNB Nutrition has finally brought us a trusted and tested form of L-BAIBA, which we call an "exercise signal" that kickstarts incredible metabolic processes! It's known as MitoBurn and it helps kick-start the 'exercise program'!
MitoBurn from NNB Nutrition is a trademarked form of L-β-Aminoisobutyric Acid, which is often referred to as L-BAIBA.
L-BAIBA belongs to the myokine family, non-protein amino acids that act like hormones. Technically speaking, hormones are messenger molecules that give instructions to the cells in your body, and that's exactly what myokines do as well.
In fact, we often refer to L-BAIBA as a muscle messenger because it gives cells the information they need to grow your muscles.
L-BAIBA in context: the exercise signal in a pill?
When you exercise, the exertion signals your body to convert the essential branched-chain amino acid, (BCAA) valine, into L-BAIBA.[22]

Leptin problem in rats? Not a problem after BAIBA treatment![23]
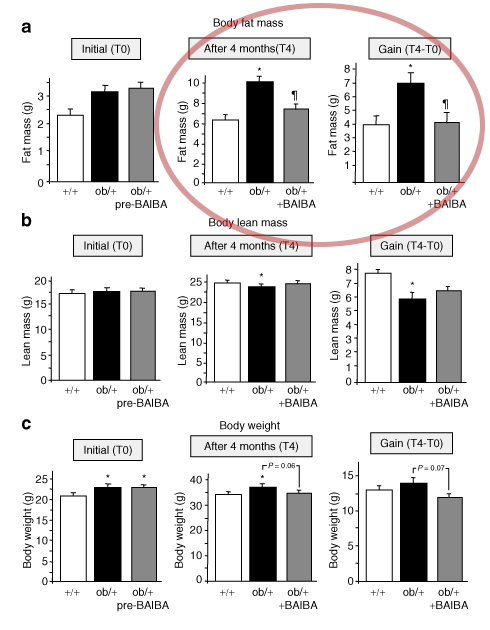
The L-BAIBA then signals to your muscle cells that exercise is occurring,[24] which in turn triggers a cascade of metabolic adaptations that help your body tolerate and recover from hardcore workouts. Examples of these changes include increased fat burning and bone protection.[22,24]
The research on L-BAIBA supplementation shows that it can:
- Increase rate of fat burning[22,23,25-27]
- Upregulate ketone production[28]
- Convert white adipose tissue (WAT) to brown adipose tissue (BAT)[26,27]
- Improve insulin sensitivity while reducing blood glucose levels[22,23,29]
- Reduce inflammation[27]
- Improve blood lipids and cholesterol[22,23]
- Increase bone density[30]
- Improve kidney function[31]
Why MitoBurn?

"BAIBA is released from the muscle after an exercise bout, promoting differentiation of brown adipocyte-like cells within subcutaneous fat depots and fat oxidation in the liver."[29]
There have been many attempts to create a good form of supplemental BAIBA, but these attempts have been fraught with difficulty.
The biggest obstacle is that only one isomer of BAIBA – the L form – does much at all to metabolism.[22,25] D-BAIBA and R-BAIBA have little effect.
That's what spurred NNB Nutrition to develop and trademark MitoBurn – it's the first pure, standardized, and stabilized form of the BAIBA L-isomer.
We wrote a whole article about this awesome ingredient. If you want to learn more, read BAIBA: Weight Loss Ingredient Generates Exercise in a Pill?! and MitoBurn: β-Aminoisobutyric Acid (L-BAIBA) from NNB Nutrition.
Why is L-BAIBA in the pre-workout support formula?
MitoBurn, as you can probably guess from the name and by reading the article we linked to, is generally used as a fat-burning ingredient. But does that always need to be the case? The answer is no.

MitoBurn (L-BAIBA) has flipped the fat burner niche on its head by supplying more of this exercise-based signaling molecule to dieters
MitoBurn is more than a "weight loss" ingredient. It's here to enhance and amplify your body's usual adaptations to exercise. By giving your body more of the information it needs to deal with intense physical exertion, you'll prime yourself to work harder and recover faster than you ever knew was possible. And this dose is no joke.
-
Citicoline - 2,000 mg
Citicoline, also known as CDP-choline, is a particularly bioavailable form of choline.
Choline's primary role in the human body is to act as a building block for the construction and maintenance of cellular membranes,[32] which are absolutely essential for cellular health and function.
Choline is also an important acetylcholine precursor, which matters because the neurotransmitter, acetylcholine, is crucial for learning and memory.[33] In fact, we at PricePlow often refer to acetylcholine as "the learning neurotransmitter" because of how important it is for memory consolidation, the process of converting short-term memories into long-term ones.
Acetylcholine upregulation translates to improved cognition in multiple domains, including learning, memory, and even psychomotor skills like balance and coordination.[34,35]
Citicoline’s special benefits
The research literature on choline indicates that citicoline, apart from other forms of choline, is particularly good at improving focus, attention, and mental energy.[36,37] It consists of two basic components, choline and uridine, both of which can improve cerebral blood flow and promote neuronal mitochondria function.[38]
Citicoline can also boost dopamine levels.[39]
All in all, it's the feel-good form of choline, included here because of its ability to improve mood, focus, and motivation.
-
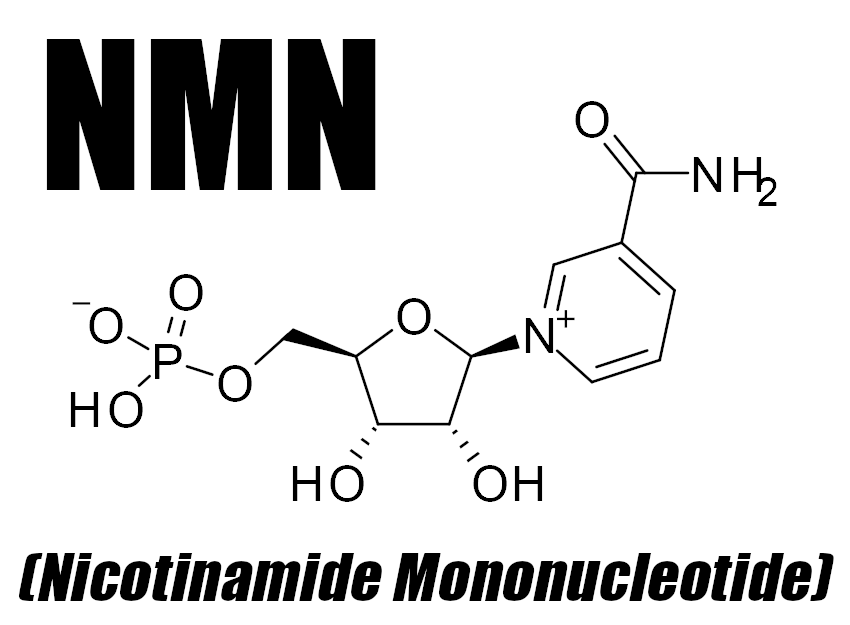
Beta-Nicotinamide Mononucleotide (as BioNMN) – 2,000 mg
BioNMN is a trademarked form of nicotinamide mononucleotide (NMN), developed by NNB Nutrition.

NNB Nutrition's BioNMN takes NMN bioavailability to a new level, and is backed by an incredibly innovative novel ingredient development team
It's pretty easy to understand why formulators would want this one in his pre-workout support formula. BioNMN can significantly increase cellular energy production by upregulating nicotinamide adenine dinucleotide (NAD+),[40] which can, in turn, lead to better performance in the gym. In fact, it's a key ingredient in two previous Formulator's Corner segments: our Mitochondria-Enhancing Pre-Workout Supplement formula and an RTD-stable Mitochondria-Boosting Pre-Workout drink.
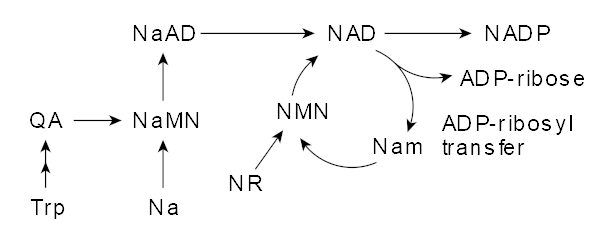
NMN: where it comes from, challenges of NMN supplementation
NMN is synthesized from niacin,[40,41] an essential B vitamin that is often touted as an energy-promoting supplement thanks to its impact on NMN and NAD+ and hence, ATP production.
The conversion pathway basically looks like this:
Niacin → NMN → NAD+ → ATP
Why is NMN better than niacin or NAD+?
You can see that niacin is an NMN precursor. We'll get some of that in below, but niacin supplementation isn't the most directly effective strategy for increasing NMN or NAD+ production.

More NMN, more food, less age-related weight gain in mice.[42] It seems that improved insulin sensitivity is a huge benefit of this mitochondrial-boosting ingredient
The main problems with niacin are that its half-life is quite short[43] and many don't want to deal with the flushing side effects of the nicotinic acid form of niacin[44] (which is ironically also added to this formula). And if you were to want the "flush-free" kind of niacin (niacinamide), doses above a certain amount can cause liver damage.[45,46]
Supplementing with NAD+ doesn't work that well either, thanks to the extraordinarily low bioavailability of oral NAD+ supplements.[47]
Fortunately, there's a better way to upregulate NAD+ and, hence, ATP: supplementing directly with NMN. It's markedly free of the issues that plague niacin and NAD+ supplementation. Its pharmacokinetic properties are much more desirable than those of niacin or its metabolites,[48] meaning it has the potential to rapidly increase NAD+ levels.
We'll dig into this as much as we can, but a full discussion of this topic is beyond the scope of this article. If you want to read more about NNB's BioNMN, check out our long-form article titled NMN Supplements (nicotinamide mononucleotide): The NAD+ Energy Precursor.
The relationship between NAD+ and adenosine triphosphate (ATP) synthesis
Let's talk for a bit about NAD+. Because NMN is a direct NAD+ precursor, upregulating NAD+ is the first step in BioNMN's ATP-upregulation process. The first thing you should know about NAD+ is that it's really important for ATP production.
Like we said earlier, ATP is your body's energy currency. It's the form of energy your cells use to carry out any and every task you ask them to perform.[41] See our "gasoline car" analogy above.

The human safety data provided in this study is great news, but even more compelling is the reduction in insulin alongside a drop in glucose.[49]
The machinery for ATP synthesis is located in your mitochondria. These "powerhouses of the cell" use a mechanism called the electron transport chain (ETC) to, in a nutshell, load chemical potential energy into the ATP molecule, which can then be liberated through hydrolysis wherever it's needed, and put to good use.
The electron transport chain works by oxidizing NADH, the chemically-reduced form of NAD+, into NAD+. The more NAD+ you have, relative to NADH, the better your ETC can work. Given the crucial importance of cellular energy for health and longevity, it's no surprise that a higher NAD+-to-NADH ratio predicts better metabolic function and a longer lifespan.[50]
Every single cell in your body is harmed by a lack of ATP-bound energy, which is why diseases of ATP production can cause serious, potentially life-threatening symptoms.[51]
Just to give one example, impaired neuronal ATP production is observed in cases of severe neurodegenerative illness.[52] That's because when there isn't enough energy to go around for all your cells, the body initiates a process called apoptosis, in which cells deliberately self-destruct in response to informational signals from the brain.[53] The purpose behind this seems to be sparing resources for other cells.

NAD Metabolism, where NMN plays a strong direct role. Image courtesy Wikimedia.
On the other hand, supplements that can upregulate ATP production are often associated with improvements in cognition[54] – cognitive health being, in our opinion, is a good proxy measure for overall metabolic health. After all, the brain is the most energy-intensive organ in the body and consumes about 20% of all the energy produced by cells despite accounting for a small proportion of total body mass.[55]
In the bigger picture, whole-body mitochondrial dysfunction has been statistically associated with insulin resistance, type 2 diabetes, and obesity,[56,57] especially when it results in the impairment of oxidative phosphorylation, the final step in producing ATP through aerobic respiration.[56]
Common-sense weight loss behaviors like eating less food and exercising more can improve insulin resistance and help reverse obesity in part because they upregulate 5' adenosine monophosphate-activated protein kinase (AMPK), an enzyme that triggers NAD+ production and activates a crucial pro-longevity protein called sirtuin-1 (SIRT1).[57]
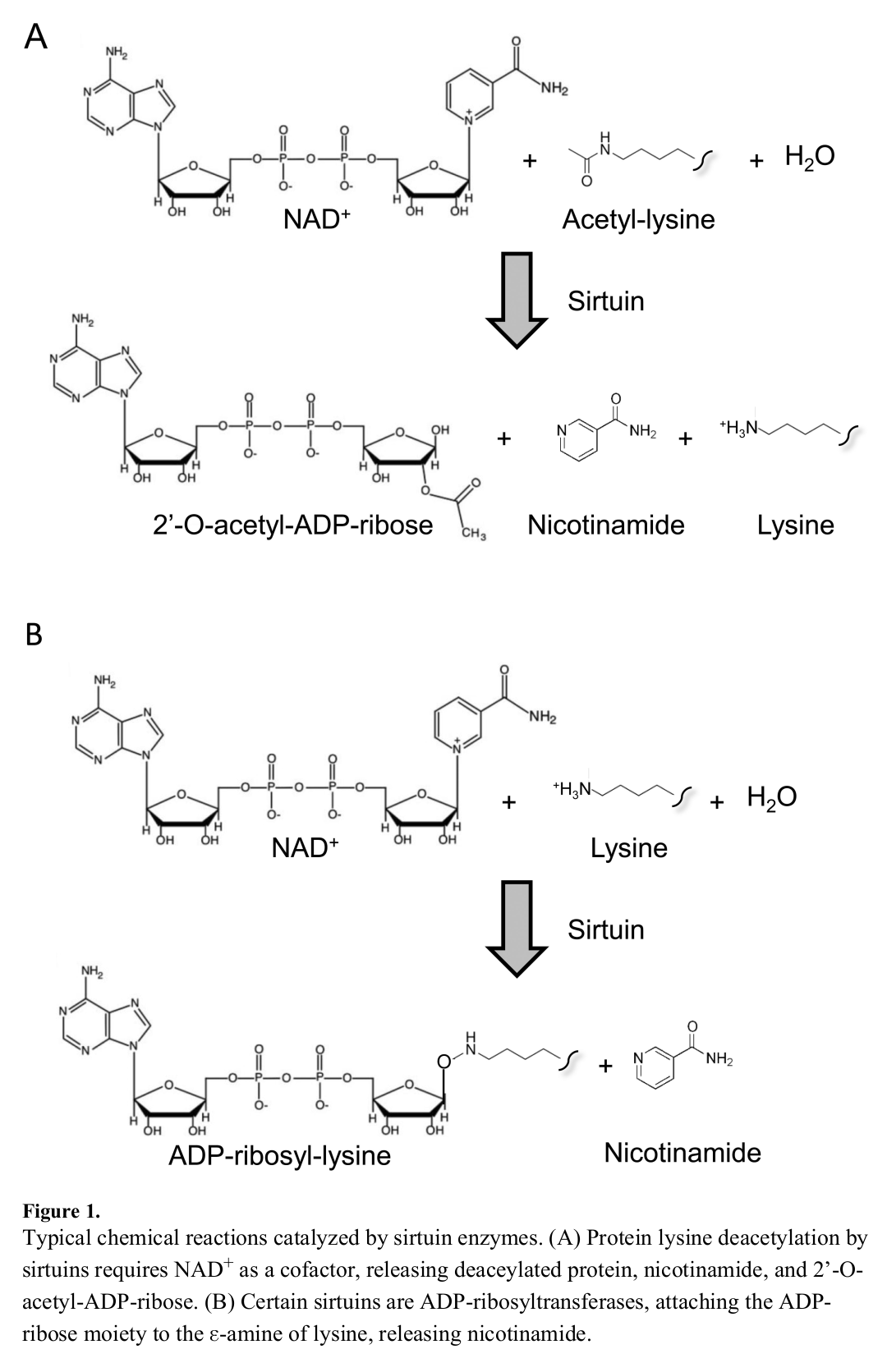
Upregulating NAD+ through NMN supplementation can activate sirtuins
One of the key benefits associated with increased NAD+ production is the ability of NAD+ to activate sirtuin proteins, which require a lot of NAD+ to function correctly.[58,59]
There are at least seven sirtuins, appropriately named sirtuins 1 through 7. That is, SIRT1, SIRT2, and so on, up to SIRT7.
Each sirtuin plays a different role in your body's metabolic function:[60]

Some reactions with respect to sirtuins and NAD[60]
- SIRT1 regulates cell growth and differentiation, apoptosis, insulin signaling, and the stress response.[60] It's also important in fighting oxidative stress,[61] which is key for optimal health and longevity.
- SIRT2 controls mitosis (the process of chromosomal duplication), and adipocyte differentiation, defends your genes against genotoxins, and helps slow the process of age-related cognitive decline.[60]
- SIRT3, SIRT4, and SIRT5 regulate the Krebs cycle, which is a crucial part of ATP production, as well as the oxidation of fatty acids (fat burning) for cellular energy, and oxidative phosphorylation (the final phase of the ATP synthesis process).[60]
- SIRT6, like SIRT2, also helps protect your genome from damage.[60]
- SIRT7 controls RNA polymerase I transcription, the process that copies ribosomal RNA.[60,62] The RNA instructions generated by RNA transcription are used by ribosomes to synthesize all the proteins used by your body, which is unbelievably important, rivaling ATP production in profundity.
Longevity research has exploded in the last couple of decades, and as the evidence mounts, it's becoming clearer each year that healthy sirtuin function is one of the primary factors regarding whether a person ages well or poorly.
NMN supplements: what the research says
OK, so now that we've defined NAD+ and why having more of it is better, the next question we need to answer is whether NMN supplementation actually increases the amount of NAD+ the body can produce. Human studies are still underway, but here's what we've said recently:
Human studies
There are a few important human studies showing benefits from NMN supplementation. These studies are primarily focused on the pro-longevity effects of NMN, which isn't exactly what we're after with this pre-workout support formula, but they can still be taken as evidence that NMN does in fact raise NAD+ levels.
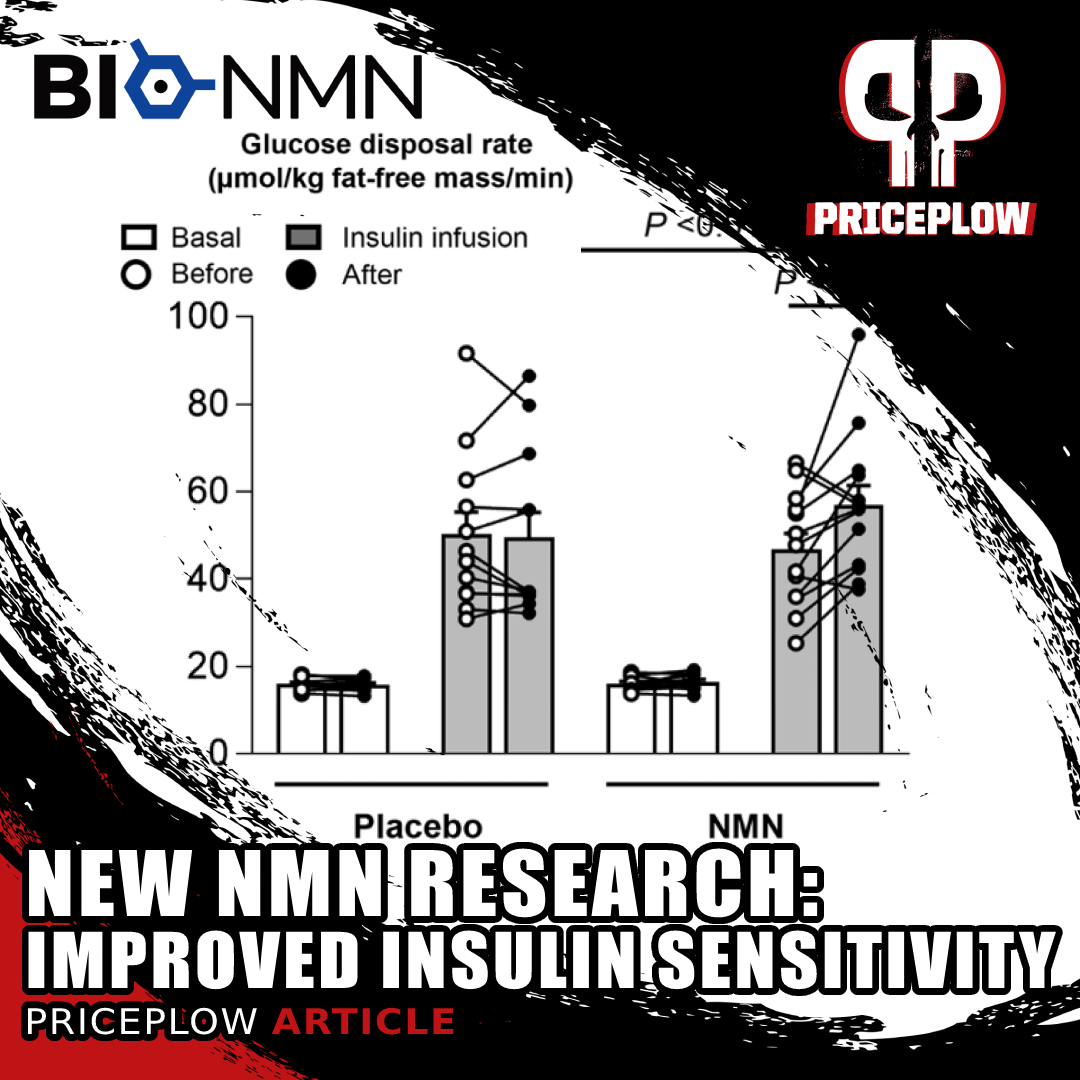
A new research study on overweight, insulin-resistant women has shown that NMN supplements improve muscle insulin sensitivity.[63]
In late 2019, the first phase of research performed by a Japaenese team was published - they were looking at acute NMN tolerance and safety. They found that doses up to 500 milligrams did not lead to any adverse effects.[49] But what's interesting is that they also noticed a strong reduction in insulin and drop in glucose within 5 hours of taking NMN![49]
In another study, 161 adults over the age of 65 were given 250 milligrams of NMN daily for 12 weeks. At the end of the study period, they had significantly lower levels of afternoon fatigue and much better leg function.[64]
Another study found that NMN is capable of increasing insulin sensitivity in prediabetic women.[63]
Finally, a randomized, double-blind, placebo-controlled study in amateur runners found that NMN supplementation helped improve their aerobic capacity,[65] which is consistent with our general observations of ATP-boosting compounds and very much a goal of this pre-workout formula.
Preclinical trials
There are additionally several animal studies showing that yes, NMN does precisely what we want it to. It has been shown to have the following properties:
Need more clean energy? Then there's a good chance you need more NAD+ -- and an incredible way to generate more is with NMN Supplementation.
- Anti-ischemic[66]
- Neuroprotective[67-69]
- Anti-diabetic[70-72]
- Anti-obesity[42,73]
- Pro-longevity[74]
All of these can be unified and explained in fundamental terms by increased cellular energy availability.
One of the animal studies we reviewed found that NMN supplementation increased NAD+ levels by 300% in aged mice. It also significantly improved certain parameters of cardiovascular function, like endothelium-dependent dilation (EDD), aortic pulse wave velocity (aPWV), and elastic modulus (EM).[74]
We've been consistently writing about ATP here on the PricePlow Blog, and one of the common denominators we see in ATP upregulation is improved arterial function through upregulated nitric oxide (NO) signaling,[75] which can obviously be beneficial to anyone who's pushing through a tough workout.
Once again, read our main NMN Supplements article to learn more. We believe this is the one ingredient in this entire list that formulators should be tinkering with in their next-generation formulas.
-
VasoDrive-AP (casein hydrolysate) – 508 mg
VasoDrive-AP is a combination of two specific milk proteins that are extracted from the casein protein fraction of whole milk: isoleucyl-prolyl-proline (IPP) and valyl-prolyl-proline (VPP).
Have an ingredient idea? Team NNB will co-develop it with you! The process is known as Ingredientology.
VasoDrive has similar effects as NO-boosting ingredients like citrulline, which is one of the industry's pre-workout mainstay ingredients we discussed earlier. However, it uses a very different mechanism of action.
Instead of triggering vasodilation by increasing NO production, VasoDrive decreases vasoconstriction by inhibiting angiotensin-converting enzyme (ACE).[76]
In terms of its overall effect on blood flow and velocity, preliminary research indicates that VasoDrive is roughly as efficacious as more traditional NO-boosting ingredients – people who supplement with VasoDrive have substantially decreased their blood pressure with effect sizes around what you'd expect from an ingredient like citrulline.[76]
Using an ACE inhibitor like VasoDrive is an awesome way to position the pre-workout support formula for synergistic effects with a conventional pre-workout.
-
InnoSlim – 500 mg
InnoSlim is a special combination of Panax notoginseng root and Astragalus membranaceus root extracts. Originally created as a weight loss support ingredient to help fight symptoms of metabolic dysfunction, it has a really interesting purpose in a pre-workout support formula.
More than ever, we understand metabolism and its critical importance. Besides exercise and cleaning up your diet from processed foods, how else can we accelerate our path back to metabolic wellness? Nuliv Science's InnoSlim is one such accelerant.
InnoSlim exerts anti-obesity effects partly by modulating the action of transporter proteins. For example, it has been shown to decrease the expression of sodium-glucose cotransporter 1 (SGLT1),[77] which is responsible for absorbing glucose from the small intestine and delivering it into the bloodstream.
Downregulating SGLT1 means less glucose absorbed from your food, which is key to InnoSlim's anti-obesity effects.
Glucose transporter 4 (GLUT4) – anabolic leverage point
InnoSlim also happens to increase the expression of a transporter called glucose transporter 4 (GLUT4),[78] which is responsible for moving glucose out of your bloodstream and into muscle tissue, where it can help drive recovery from exercise.
When you put this together with the fact that InnoSlim can activate certain metabolic pathways that are involved in fat and glucose metabolism, like AMPK, ACC-P HIF-1, and adiponectin,[79] it's clear why InnoSlim might belong in a pre-workout support formula.
Through activating GLUT4, InnoSlim primes your body to properly dispose of glucose consumed post-workout.
It can also, through adiponectin activation, prevent glucose from getting stored in your liver, which is a boon both for skeletal muscle-glycogen replenishment and long-term metabolic health.[80]
-
Niacin (as nicotinic acid) - 250 mg
Anyone following along in the PricePlow Discord knows that we occasionally get carried away with the niacin flush, fondly known as the "floosh".
Regardless of the B vitamin's limitations discussed above in the BioNMN section, giving your body an adequate supply of niacin (Vitamin B3) is still excellent for supporting healthy cellular energy production.

For NAD+ production, we believe that NMN > Niacin... but if you're okay with the "floosh", why not use both?
For one thing, niacin has been shown to upregulate adiponectin,[81] a hormone we also discussed in the section about InnoSlim. It has a multitude of effects on human metabolism, chief among which is a significant increase in insulin sensitivity.[82]
Adiponectin also activates AMPK,[83] which again, is crucial for maintaining ATP balance.
Niacin reduces the amount of inflammation caused by extra body fat,[81] which is great because this particular kind of inflammation is closely associated with metabolic disease.[61] This mechanism may not acutely work for pre-workout purposes, but in the long run, consistent niacin supplementation can improve overall metabolic function, which can support harder training.
Niacin and exercise
Some studies have found that niacin can actually improve athletic endurance, in part because a steady supply of surplus niacin can upregulate fast twitch muscle fibers that are more fatigue-resistant than other types of muscle fiber.[84] In the same study, the researchers speculate that niacin may have helped drive free fatty acids into muscle tissue, which in turn utilized those FFAs as a substrate for cellular energy production.[84]
What seems more certain is that animals treated with niacin are better able to utilize glucose for fuel than animals treated with a placebo.[84]
Note that we've of course chosen the flushing type of niacin - nicotinic acid - in this formula. 250 milligrams generally provides a low amount of flushing - the flushing gets far more pronounced at around 500 milligrams.
-
NeuroFactor Whole Coffee Fruit Extract - 200 mg
NeuroFactor is an extract of whole coffee fruit designed to improve cognitive function, and particularly, focus.
We were tasked to make a mitochondrial health boosting pre workout supplement that provided clean energy. Inside is quite the formula that we'd love to try out!
Its primary mechanism of action is increasing the expression of brain-derived neurotrophic factor (BDNF), a protein that supports the growth, survival, and maintenance of neurons in particular regions of the adult brain.[85]
Scientists have identified a significant positive correlation between BDNF blood levels and broad cognitive performance.[86-89] It appears that higher levels of BDNF, especially later in life, may protect the aging brain from toxic insults and neurodegenerative illnesses.[86,90]
On the other hand, having low BDNF levels seems to raise one's risk for neurodegenerative illness.[90]
BDNF is also important in the short-term for cognitive performance, and has actually been proposed as the primary mechanism behind near-miraculous improvements that aerobic exercise confers on brain function.[87]
That isn't surprising, since one of BDNF's key roles is to regulate neuroplasticity, the ability of the brain to physically remodel itself in response to learning. In particular, BDNF is crucial for neuroplasticity in the hippocampus region of the brain, where a process called long-term potentiation (LTP) occurs.[88] By supporting LTP, BDNF can help the brain convert short-term memories into long-term memories, which is obviously crucial for acquiring new skills or information.
Research on NeuroFactor indicates that a 100-milligram dose is enough to increase BDNF blood levels by almost 150%.[90] Twice the dose isn't necessarily twice as good, but in this formula, we obviously go above and beyond.
-
Senactiv (Panax notoginseng (root) and Rosa roxburghii Tratt (fruit) Extracts) – 50 mg
Senactiv from NuLiv Science is a senolytic agent, which is defined as a drug or compound that helps remove old, dead, or dying cells to make way for newer, more functional and healthy ones.

Improved VO2 Max from Senactiv means big things for everyone, not just endurance athletes! Image courtesy NuLiv Science.
To date there have been four human trials and two in-vivo studies on Senactiv's bioactive constituents.[91-96] One found that people supplementing with Senactiv's ginseng constituent an hour before exercise showed signs of reduced muscle damage, accelerated glycogen replenishment, and decreased inflammation following the completion of their workout.
One of the important mechanisms behind Senactiv's ability to limit exercise-induced damage is its ability to inhibit lipid peroxidation.[91] This is important because the toxic aldehydes formed by the peroxidation of lipid membranes are a huge contributor to the damage caused by oxidative stress throughout the body.[97]
But the really interesting thing to note about Senactiv's role as a pre-workout ingredient is that in one study, it increased time to exhaustion at 80% VO2max by about 20%.[91] That's a substantial boost to athletic endurance.
-
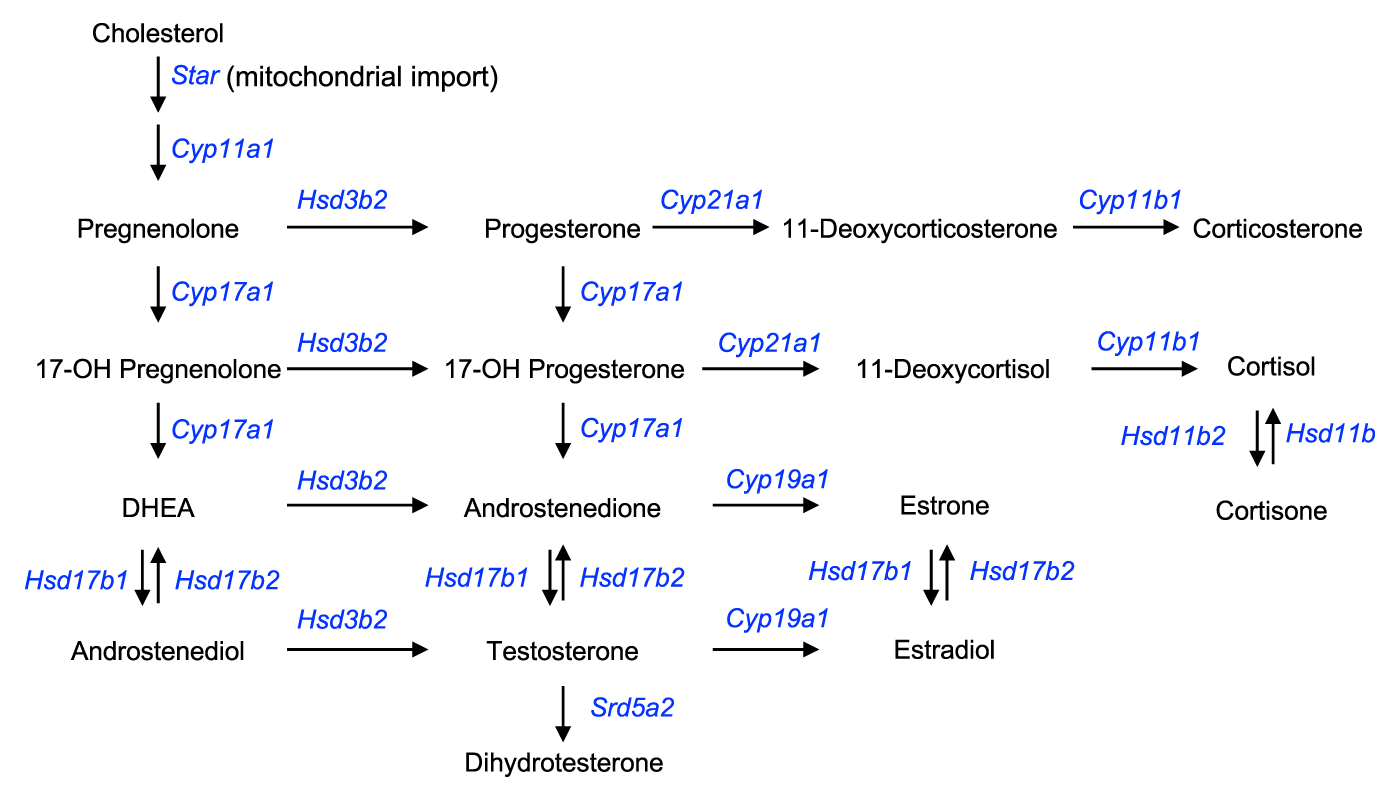
Pregnenolone – 100 mg
Pregnenolone is the most important hormone you've never heard of.
It's the precursor to all of your body's steroid hormones – including testosterone and its metabolites.
We say precursor because although there are some intermediate steps in the steroid hormone synthesis pathway, it remains the case that all steroid hormones begin life as a pregnenolone. Every single molecule of testosterone in your body was pregnenolone first.

The steroid synthesis pathway – all hormones on this chart are converted from pregnenolone through a series of enzymatic reactions.[98]
Although exercise can be extremely beneficial when performed at the right level of intensity and frequency, it is possible to overdo exercise because, at the end of the day, it is a stressor. You get stronger, faster, and more fit by compensating for the stress that exercise induces on your body – this is what we mean when we talk about "recovery from exercise."
Exercising too much, and at too high of an intensity level, can lead to a syndrome called overtraining, which most of us have heard about before.
But problems can occur on the other side of the equation as well. If your capacity to recover from exercise is impaired, your normal exercise regimen might cause health issues for you.
Specifically, exercise has been shown to transiently elevate stress hormones like adrenaline and cortisol. If, for any reason, you lose your capacity to bring these hormones back into balance with antagonistic youth-promoting hormones like progesterone, testosterone, and DHEA, you can end up stuck in a state of chronic stress. In other words, you can experience overtraining syndrome, even if your volume of training is not extraordinary.
Your body only makes so much pregnenolone at once, so if cortisol pathways are chronically upregulated, that will deplete your body's supply of cortisol at the expense of other, more beneficial steroid hormones.
Supplementing with extra pregnenolone can help normalize your steroid hormone synthesis pathways, bringing your steroid-hormone profile back into balance.
This has been validated in multiple domains of medical research. Even in cases of extreme mental illness, supplemental pregnenolone has been shown over and over again to bring cortisol levels down and restore hormonal balance.
So, long story short, pregnenolone is here to help your body cope with the long-term stress associated with completing an intense exercise and/or dieting regimen. Or, at the very least, it gives you a wider margin of error when figuring out what kinds of demands your body can handle.
-
Methylcobalamin – 1 mg (41,667% DV)
We always appreciate seeing the methylcobalamin form of vitamin B-12 because, compared to its other form -- cyanocobalamin -- this one is far superior in terms of bioavailability and methylation status.
The presence of the methyl group is a huge reason why methylcobalamin is preferred over cyanocobalamin. When taken in large doses, methylcobalamin can act as a methyl donor, affecting cellular energy production and metabolic processes all over the body in ways that go far beyond merely satisfying vitamin B-12 requirements.
One great example of this is the methionine-homocysteine cycle, a process in which homocysteine is methylated (bonded to a methyl group) and converted into methionine, an essential amino acid. This is a crucially important process because homocysteine overload is linked to chronic inflammation and cardiovascular disease. It can even develop into cognitive dysfunction.[99]
Don't be put off by the huge dose – B-12 is a water soluble vitamin, so your body should safely eliminate any excess you consume.
NNB Nutrition is now offering brands Whitelabeled Gummies of their novel ingredients like MitoPrime (L-Ergothioneine) and RhodioPrime 6X -- and they are superb!
Anecdotally, many people report a boost in energy after taking a B-12 megadose, which is one reason why B-12 injections have been favored as an energy-boosting supplement by luminaries like former UK prime minister, Margaret Thatcher.[100]
Note that in Joey's original spreadsheet, he has 100 milligrams of material, but that yields 1 milligram of the actual vitamin.
-
5-hydroxytryptophan (5-HTP) – 25 mg
You've probably heard of 5-HTP as a serotonin precursor, and supporting optimal neurotransmitter production is usually a good thing.
But did you know that 5-HTP might improve body composition as well?
At least one study has found that eight weeks of supplementation with 100 milligrams of 5-HTP can favorably alter body composition by encouraging fat loss.[101]
5-HTP can also potentially help you sleep: it's an important intermediary step in melatonin synthesis. The process goes like this:[102]
- L-tryptophan is converted into 5-HTP
- 5-HTP is converted into serotonin through a decarboxylase enzyme
- Serotonin is converted into N-acetyl-serotonin by the enzyme hydroxyindole-O-methyltransferase
- N-acetyl-serotonin is converted to melatonin[102]
This ingredient shouldn't put you to sleep, but rather, provide your body with the raw materials needed to synthesize melatonin at the appropriate time.
-
Pyrroloquinoline Quinone (PQQ) – 20 mg
Once again, sleep quality and stress management are crucial to maximizing gains from exercise, and maintaining optimal health throughout a demanding exercise regimen.
Tired of pre-workout drinks with too much caffeine? Using NNB Nutrition's novel ingredients, we formulated a hypothetical mitochondrial health boosting pre workout RTD in Formulator's Corner #08!
Pyrroloquinoline quinone (PQQ) is a special salt that can drive mitochondrial biogenesis,[103,104] the process by which your body can create new mitochondria. It does this by activating a key sirtuin, SIRT1, which we discussed in the section on NNB Nutrition's BioNMN![103]
One interesting effect of increasing mitochondrial density and, hence, increasing ATP production is improved blood flow and oxygen uptake.[105]
In the brain, this can look like better cognitive performance,[106] but can also translate over time to lower perceived stress levels, higher energy levels, and improved sleep.[107]
And of course, as we have discussed in previous sections, optimizing ATP production is a huge potential benefit for athletic performance and recovery.
We're a huge fan of PQQ as a mitochondria-optimizing ingredient, and glad to see it getting more support here. It's also in our theoretical Formulator's Corner Mitochondria-Boosting Pre-Workout supplement and accompanying RTD. We hope the industry will use it more frequently going forward.
-
Vitamin B6 (as Pyridoxal-5-Phosphate) – 10 mg (590% DV)
Vitamin B6 is an important factor for coenzyme production, which in turn affects tons of metabolic processes and cellular energy availability.
Just to give two examples, your body needs B6 to make pyridoxal 5' phosphate (PLP) and pyridoxamine 5' phosphate (PMP), two enzymes that together influence several hundred different metabolic processes in your body.[108] Among the most important of these processes is protein metabolism, for which PLP and PMP are absolutely essential.[108]
For the sixth installment of our Formulator's Corner series, we've drafted up a powerful Immunity Supplement with modern concerns in mind -- and it features NNB Nutrition's MitoPrime (L-Ergothioneine)
B6 is also an underrated neurotransmitter precursor. And, as we've discussed, neurotransmitter levels can have a pretty significant impact on mood and motivation, two things that will make your workout much easier if they're dialed in.
Another important process controlled by B6 is homocysteine regulation.[108] Since high homocysteine levels can be a major factor in the onset of cardiovascular disease, you definitely want to give your body everything it needs to keep homocysteine under control – that means B6.
Vitamin B6 is also important for hemoglobin production,[108] which is obviously important for athletic performance.
It can also affect immune function, gluconeogenesis, and glycogenolysis (the degradation of glycogen for usable energy).[108]
Supplementation with B6 is statistically associated with lower risk of cardiovascular disease, cancer, and neurodegenerative illness.[108]
-
Riboflavin – 10 mg (769% DV)
Also known as vitamin B2, riboflavin is implicated in a few crucial metabolic processes, including the electron transport chain (ETC) and fat metabolism.[109]
ETC is what produces ATP,[109] so riboflavin is obviously important for whole-body energy production, which is why we're seeing it show up here.
-
Sebrium DCD (std. to >3% mesembrine) – 5 mg
Mesembrine is an alkaloid derived from the kanna plant (Sceletium tortuosum).[110] It has significant psychoactive effects.
The primary mechanism of action behind mesembrine is serotonin upregulation,[111] which can give the user a big boost in mood and perceived energy.
The source of mesembrine, S. tortuosum, has a long history of use in folk medicine for the management of emotional disorders, and has even been investigated as a potential treatment in modern neuropharmacopsychiatry.[112]
The idea here is, again, to give you the focus, energy, and motivation to make your workout feel as pleasurable and effortless as possible.
-
Arcofolin (folate) – 5 mg (1250% DV)
For our second Formulator's Corner, we've envisioned a very thermogenic fat burner with four premium ingredients from NNB Nutrition, alongside two of our favorite stimulants
Folate (vitamin B9) is the general name for a number of chemical compounds that structurally resemble folic acid.[113] Good whole-food sources of folate include egg yolks, oranges, lemons, beans, and leafy green vegetables like spinach.[113]
Folate is an essential vitamin, meaning that your body can't make it, and you must obtain it from your diet.[113,114]
Folate is used for DNA synthesis and repair, cellular replication, maturation of red blood cells, and the homocysteine-methionine cycle, which we've discussed in previous sections.[114]
Folate deficiency can lead to anemia, birth defects, arterial disease, and cancer, and is definitely something that should be avoided.[114]
Arcofolin is a trademarked form of methylated folate. Just like with the methylated Vitamin B6 and B12 additions, we like seeing this form given the crucial role that methylation plays in human metabolism.











Conclusion

Glaxon NAD+ Synergy uses BioNMN (NNB Nutrition's nicotinamide mononucleotide ingredient) combined with two synergistic ingredients to keep NAD+ levels high for incredible cellular energy!
This was a fun thought experiment – we're super stoked to see such high doses of BioNMN and MitoBurn from NNB Nutrition. These are two of our favorite ingredients, so we love having the opportunity to write about them.
Of all of the above ingredients, BioNMN is the one that we'd love to see in more pre-workouts, and PQQ could make a unique non-stimulatory addition as well.
Additionally, the inclusion of pregnenolone is also super interesting, and an avenue that we hope the industry will consider for further exploration and development.
Stay tuned for future Formulator's Corner segments, and thanks to Joey and Dave Cook for putting this one together!

Comments and Discussion (Powered by the PricePlow Forum)