In our first edition of Formulator's Corner, we proposed a brain-boosting nootropic coffee creamer that used multiple ingredients to increase the mental power of a cup of coffee. Today we're back with another idea. This time, we turn our attention to fat-burning.
While nootropics have seen rapid growth in the supplement industry over the past few years, fat burners and other dieting aids are a foundational category of products. Losing weight and reducing body fat are common sense goals — less body fat, improved metabolic health, and overall wellness are appealing to everyone. Though this well-trodden path is best traveled by effective training and proper nutrition, fat burners and metabolic-boosting products can unlock additional gains.

For our second Formulator's Corner, we've envisioned a very thermogenic fat burner with four premium ingredients from NNB Nutrition, alongside two of our favorite stimulants
Sports supplement companies have recognized this demand and have done an excellent job satisfying it. Fat-burning products are consistently among the most popular dietary supplements available. According to the National Institute of Health, Americans spend roughly $2.1 billion annually on weight-loss supplements in pill form[1] — and that figure doesn't even account for powders, liquids, and bars!
Whether delivered in a powder, a few capsules, or a ready-to-drink liquid, brands have created many well-formulated weight-loss aids. Some leverage stimulants, such as caffeine, to ignite thermogenesis and provide energy, while others refrain from stimulant use and instead opt for herbs and other ingredients to improve metabolic functioning. Many products combine the three, creating effective formulas that help influence weight and body fat on multiple fronts.
Our Take: Fat Loss Through Multiple Premium Ingredients
Considering that there are plenty of great fat burners already out there, we recognize the need to do things a little differently to make our formula stand out. To do that, we're relying on our friends over at NNB Nutrition, a leading ingredient formulator and innovator.
NNB Nutrition is an innovative ingredient development company with an elite team of over 100 scientists from over 10 countries, and some incredible metabolic aids.
Many fat-burning formulas contain a mix of ingredients — some are simple additions, while others are patented, standardized extracts. Standardized inclusions are premium ingredients, and can admittedly drive up the cost of a supplement if too many are used. Thus, most weight-loss aids will rely on a combination of lower- and higher-cost ingredients to get the job done. While this is advantageous from a business standpoint, since we're only spitballing here, we decided to shake things up a bit. What about a fat burner that almost exclusively uses higher-cost, premium ingredients supported by clinical data?
Our fat-burning formula is heavily driven by four premium NNB Nutrition ingredients, which alongside two additional stimulants, deliver potent and comprehensive weight-loss benefits. This formula sets itself apart from other fat burners by being composed almost exclusively of high-quality, cutting-edge ingredients that give it an incredibly versatile profile - and one of them hasn't even been used in a fat burner yet, so it's guaranteed to offer something unique!
Team PricePlow ultimately settled on six ingredients to use in our formula, four of which come directly from NNB's industry-leading labs:

Brands are looking for unique formulas that actually work. We believe we've got one here, but it will admittedly cost a bit more than the average weight loss aid!
- NNB Nutrition's MitoBurn, a biologically active form of BAIBA that ignites the metabolism, influencing multiple metabolic factors while also priming stored fat tissue for expenditure.
- ZinjaBurn, NNB Nutrition's ground-breaking dehydrozingerone extract that delivers the metabolic effects of curcumin while circumventing potential pitfalls in bioavailability.
- Caffeine anhydrous, the tried-and-true energy base that also possesses thermogenic capabilities.
- GlucoVantage, NNB Nutrition's dihydroberberine ingredient that optimizes blood sugar levels and insulin sensitivity.
- Grains of paradise in the form of CaloriBurn from NNB Nutrition. This spicy inclusion — standardized for its bioactive constituents — turns up the heat, activating fat tissue for metabolization.
- Synephrine from bitter orange extract, which further revs the metabolic engine and works in synergy with caffeine.
Below, we'll discuss what each of these ingredients brings to the table, and why, together, they form an excellent fat-burning product. But first, it's important that we mention that many of these ingredients have uses in areas other than weight loss and improved metabolic health. For the purposes of this post, though, we're going to focus solely on their benefits in terms of weight and metabolic functioning.
![]()
Our Fat Burner Ingredients
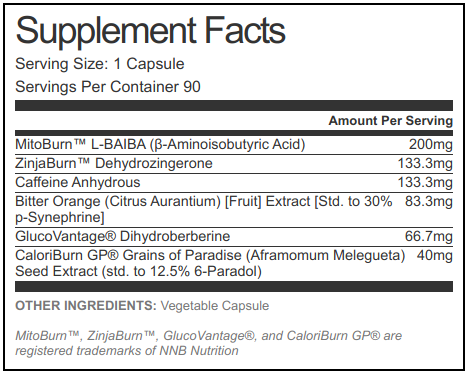
Our NNB powered fat burner brings 400mg caffeine per day at the max dosage, alongside some potent thermogenics and blood sugar modulators
Our formula is designed for three capsules daily, which we'd suggest being taken in the following fashion: two capsules in the morning, one capsule in the mid-afternoon (depending on stimulant tolerance). This protocol is flexible, of course — some individuals may prefer a different timing scheme, others may prefer not splitting doses at all, and others may only decide to take one or two capsules per day. The doses we've decided to use are all assuming a daily three capsules, though we've listed per capsule amounts, too.
Here is what you get in our NNB Nutrition-driven fat-burning supplement:
-
MitoBurn (L-ꞵ-Aminoisobutyric Acid) - 600mg (200mg per capsule)
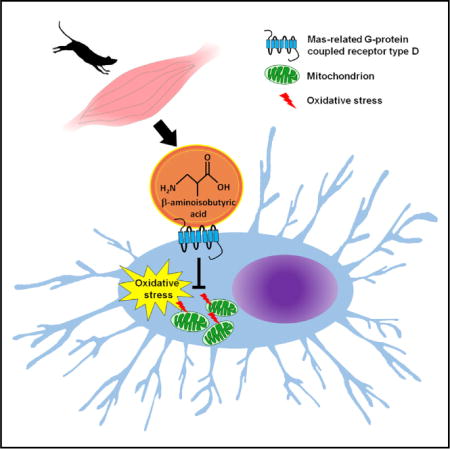
ꞵ-aminoisobutyric acid, also known as BAIBA, is an amino acid used by the body as a signaling molecule. Formed through the breakdown of either valine or thymine, BAIBA is generated during muscle contraction — specifically, this myokine is released from skeletal muscle in response to exercise stimulus.[2] Though this amino acid isn't used to build muscle (like the nine essential amino acids), BAIBA does helps initiate various biological responses of exercise, mainly those related to metabolism.
BAIBA signals PPAR

NNB Nutrition has finally brought us a trusted and tested form of L-BAIBA, which we call an "exercise signal" that kickstarts incredible metabolic processes! It's known as MitoBurn and it helps kick-start the 'exercise program'!
In response to an exercise stimulus, the skeletal muscle initiates a chain of reactions that ultimately lead to what we hope that stimulus will yield — muscle repair and growth and increased thermogenesis. These responses begin with the peroxisome proliferator-activated receptor-γ coactivator-1⍺ (PGC-1⍺). PGC-1⍺ is a transcriptional protein that spikes in activity in skeletal muscle during and after aerobic activity.[3] PGC-1⍺ activity is related to various mechanisms that are beneficial for our health (such as mitochondrial biogenesis, improved insulin sensitivity, fatty acid oxidation, thermogenesis) by coactivating multiple transcription factors and nuclear receptors throughout the body.[3]
PGC-1⍺ operates mainly within myocytes, however, and cannot traverse across the body itself. Instead, this protein uses myokines to communicate its upregulation, with irisin and BAIBA doing its bidding elsewhere. BAIBA signals peroxisome proliferator-activated receptor-γ (PPAR-γ) and peroxisome proliferator-activated receptor-⍺ (PPAR-⍺), which mediate fatty acid metabolism and adipocyte differentiation.[2,4,5] BAIBA upregulates the activity of uncoupling protein-1 (UCP-1) and cell death-inducing DFFA-like effector a (CIDEA), two genes unique to brown adipocytes. These fat cells form brown adipose tissue (BAT), which is the energy-spending variant of fatty tissue that the body holds.
Brown adipose tissue (BAT) vs white adipose tissue (WAT)
The body consists of two major types of adipose tissue — brown (BAT) and white (WAT) — whose purposes differ significantly:
- WAT makes up the majority of fat the body holds, storing excess energy as the visceral fat that we can see and grab on our person. WAT also creates leptin, the hormone that communicates satiety and signs of energy balance to the brain.[6] WAT stores can be tapped into for expenditure during periods of caloric restriction and low-intensity activity.
- BAT represents the fatty tissue used for thermogenesis.[6] BAT houses brown adipocytes, which stimulate respiratory chain activity, and thus oxidation, through UCP-1 activation. Though the body only holds small amounts of BAT, a higher BAT-to-WAT ratio raises the potential to use fatty acid stores for energy.
PPAR-induced WAT “Browning”
BAIBA increases the activity of UCP-1 and CIDEA, two genes that allow brown fat cells to be oxidized and used for thermogenesis. According to research published in Cell Metabolism in 2014, BAIBA activates these genes in white adipocytes via PPAR-⍺ activation,[2] effectively "browning" WAT into "beige" fat,[2,6] which allows for the use of stored fat for heat generation. In other words, BAIBA facilitates the burning of stored body fat by giving it the necessary characteristics for oxidation.
Fatty acid oxidation via BAIBA doesn't just end there, either. PPAR-⍺ activation also increases the release of fatty acids from the liver for even more fat-burning,[7] in addition to optimizing cholesterol levels in the process.[8]
Other BAIBA-mediated processes
For obvious reasons, BAIBA's ability to free up more body fat for burning grabs much of the spotlight. However, this signaling amino acid is capable of much more in regards to improving metabolic health, too:
Enhanced ketone production
Beta-hydroxybutyrate (BHB) tends to be discussed mainly in relation to ketosis and the ketogenic diet. And while this energy-producing ketone is integral to both of these concepts, it's still important in non-ketogenic situations. BHB is produced in the liver during fatty acid oxidation,[9] an important note considering the link between hepatic fat oxidation and overall metabolic health.[10] BHB helps facilitate the burning of fat in the liver such that fat stores elsewhere can be used for energy. This has relevance for any dieter, not just someone in ketosis.

Beta-aminoisobutyric acid (BAIBA), induces beneficial effects on lipid homeostasis in mice.[11]
In a 2003 study published in Antimicrobial Agents and Chemotherapy, researchers tested the effects of BAIBA on mitochondrial and metabolic processes in mice. In a placebo-controlled manner, mice were either fed 10 milligrams per kilogram of body weight of BAIBA daily or 100 milligrams per kilogram of body weight of BAIBA daily for two weeks. They found that the 100-milligram group saw a 64% increase in total ketone bodies, reflecting enhanced mitochondrial beta-oxidation of fatty acids in the liver.[12]
One year later, a study published in Antiviral Therapy reached similar conclusions. They found that BAIBA supplementation increased plasma BHB levels in lean mice and led the team to hypothesize superior hepatic fat-burning that would ultimately lead to reductions in overall fat mass.[13] Sure enough, further tests confirmed their beliefs — despite zero changes in diet, these mice saw significant decreases in body fat mass compared to baseline.[13]
BAIBA encourages the production of BHB, the primary ketone body formed when oxidizing fat in the liver. This helps clear out hepatic fat stores, allowing fat located elsewhere to be used for energy. If you're looking to shed a few pounds, this uptick in BHB production can be incredibly useful, even more so if you're a keto dieter.
Warding off leptin-related issues
As mentioned earlier, leptin is the hormone that controls satiety and is produced in WAT — the more body fat one holds, the more leptin one likely has. However, research has shown that though obese individuals see higher leptin production, they have decreased leptin sensitivity, which limits their ability to feel full and satiated.[14] This encourages more energy consumption, further tipping the scales (literally) in the favor of weight gain.
In a study published in a 2012 edition of Obesity, researchers tested two different doses — either 100 or 500 milligrams per kilogram body weight per day — of BAIBA in leptin-deficient mice for four months. In addition to treatment, these mice were also fed a high-calorie diet. Researchers found that BAIBA fully prevented body fat gain, as well as other markers of obesity like glucose intolerance.[11] They cited such effects not only to increased plasma BHB levels but also to leptin stimulation.[11]

Despite eating the most calories, the high-fat diet mice that also received BAIBA had far less weight gain and better blood glucose scores than their non-BAIBA counterparts.[15]
Though these improvements in leptin sensitivity and body fat levels were not seen in mice that were completely deficient in the satiety-regulating hormone, the above results are promising nonetheless. Warding off leptin deficiency or leptin intolerance can help attenuate fat mass gain and aid in the process of losing unwanted fat.
Improved insulin sensitivity
As mentioned above, glucose intolerance is a key marker of poor metabolic health and is tied to many diseases and conditions. Insulin resistance is an issue seen often in obesity, let alone glucose-specific conditions like diabetes.
BAIBA is capable of improving glucose tolerance in both cases. In a 2015 study published in Diabetologia, researchers found that BAIBA supplementation attenuated the development of glucose intolerance in mice being fed a high-fat diet.[15] Another study, found in a 2016 edition of Scientific Reports, saw BAIBA improve insulin sensitivity in mice with type 2 diabetes.[16]
These findings suggest that BAIBA is capable of maintaining healthy insulin sensitivity and improving it in cases in which it's already damaged. Along with everything else this compound has shown, it looks like we have a full-fledged metabolic-improving ingredient on our hands.
Safety - an important note
It's imperative that we reiterate that there's little human research out there, making it difficult to fully assess the safety of BAIBA supplementation. However, a 2019 third-party analysis provided by NNB Nutrition saw no mortality or adverse effects in doses as high as 2,000 milligrams per kilogram body weight in mice.[17] This suggests that BAIBA has a relatively high tolerance and can be safely supplemented with, as these doses convert to as much as 1.5 grams per day for a 220-pound human. However, the data supporting the effects of this ingredient have not been fully evaluated by the FDA, and as such, one should consider potential usage with the utmost care and caution.
MitoBurn - NNB Nutrition's bioactive L-BAIBA ingredient

New research has shown that BAIBA may protect from disuse-based bone loss thanks to it preventing osteocyte cell death that would have been induced by reactive oxygen species (ROS).[18]
When research unveils a potent ingredient that has the potential to break new ground in supplementation, NNB Nutrition is often leading the charge of bringing it to market. This is evidenced by their BAIBA variant MitoBurn, a ground-breaking fat-burning ingredient that we're opting for in our formula.
MitoBurn is a high-quality variant of L-BAIBA, which is the biologically superior form of BAIBA. BAIBA comes in two forms — L-BAIBA and D-BAIBA. The former is a product of valine metabolization, while the latter comes from thymine breakdown.[18,19] Research has shown that L-BAIBA is the variant found in plasma as a result of muscle contractions, and is likely the form of BAIBA related to the signaling amino's metabolic effects.[18,19]
NNB Nutrition created MitoBurn with this knowledge in mind, developing an L-BAIBA ingredient that is biologically active and HPLC-tested for purity. This formulation packs the comprehensive metabolic punch shown in clinical data, igniting the metabolism and fatty acid oxidation for efficient, effective fat loss.
Dosage
Using existing research, as well as the safety assessment mentioned earlier, NNB Nutrition recommends dosing MitoBurn at 250 to 500 milligrams, taken one to two times per day. In our fat burner, we're operating in the middle-ground of this recommendation, using a total of 600 milligrams split evenly between three pills. Our protocol has 400 milligrams coming in the morning followed by 200 milligrams midday. This flips the metabolic switch for effective fat-burning early, and encourages continued thermogenesis and fat oxidation later on in the day.
To learn more, read our articles titled BAIBA: New Weight Loss Ingredient Generates Exercise in a Pill?! and MitoBurn: β-Aminoisobutyric Acid (L-BAIBA) from NNB Nutrition.
-
ZinjaBurn (Dehydrozingerone) - 400mg (133.3mg per capsule)
Various natural herbs and roots with histories in folk medicine are often seen in supplementation. Many of these ingredients are potent antioxidants that bring along a few other benefits. One of these natural ingredients is ginger, commonly leveraged for its three phenols — gingerol, shogaol, and zingerone — that have biological capabilities in the body.

Meet ZinjaBurn from NNB Nutrition. With a similar structure to curcumin, it has anti-inflammatory effects and better bioavailability.. but its effects skew towards weight loss related benefits!
These phenols are believed to be the drivers behind ginger's uses in relation to diet and weight. In a 2012 study published in Metabolism, administration of ginger alongside a high-carbohydrate meal increased energy expenditure in males.[20] And because 6-gingerol has been shown to activate PPAR-⍺ — increasing fatty acid oxidation via the "browning" of white fat — it is often awarded credit for the herb's metabolic effects.[21,22] When it comes to metabolic-boosting, fat-burning potential, however, one of ginger's other phenols, zingerone, deserves its fair shine, too.
Dehydrozingerone (DHZ), a supplemental form of zingerone, is a half analog of curcumin, the main bioactive metabolite in turmeric (a root belonging to the same family of plants as ginger). Curcumin is often used for its antioxidant and anti-inflammatory capabilities, though it also has potential as a metabolic aid. Such potential is difficult to yield in practice, however, due to extremely poor bioavailability.[23] DHZ circumvents this, as the ginger-based constituent holds superior bioavailability compared to curcumin and is more effective in supplementation.[24] Such effects are rather potent in terms of improving metabolic health, making it an excellent addition to our formula.
Metabolic boost via AMPK stimulation
DHZ's action in the body is what warrants both its accreditation in the potency of ginger supplementation and its use in place of curucmin for metabolic health.
Adenosine monophosphate kinase (AMPK) is an enzyme that regulates carbohydrate and lipid metabolism.[25] Activated in skeletal muscle during exercise, AMPK stimulates energy-generating processes that demand adenosine triphosphate (ATP) such as glucose uptake and fatty acid oxidation, while simultaneously suppressing energy-storing processes like lipid storage.[25] In other words, AMPK activation ignites the metabolism, revving the body's engine and increasing its demand for energy. This encourages caloric expenditure, which in terms of weight loss, encourages fat loss.
In a 2015 study published in the Journal of Cellular and Molecular Medicine, researchers assessed the metabolic effects of DHZ on mice fed a high-fat, hypercaloric diet in a placebo-controlled experiment. They found that DHZ suppressed weight gain and lipid accumulation due to AMPK stimulation.[24] Effectively maintaining an energy-demanding environment, DHZ attenuated the energy storage that comes with a hypercaloric diet.
Improves insulin sensitivity
Now that we've covered the incredible effects of dehydrozingerone, it's time to talk about where to get it!
The researchers also assessed various other markers of metabolic health, specifically those related to AMPK stimulation. One such marker is insulin sensitivity — the body's ability to transport glucose effectively and reduce blood sugar is improved with more efficient AMPK activity. This 2015 study found that the DHZ-induced AMPK stimulation increased GLUT4 activity.[24] GLUT4 is the main transporter that moves glucose through the body, and therefore its heightened activity bodes well for blood sugar levels and overall metabolic health.
Sure enough, GLUT4 activation yielded conclusive results in this regard. They found that the mice fed DHZ saw a 35% reduction in blood glucose levels and a 30% reduction in insulin activity.[24] Considering that improved glucose uptake and insulin sensitivity are positively related to reductions in weight and fat gain,[26] this action holds significant potential for dieters.
Better than curcumin for diet applications!
Though the above study uses mice, such effects have been seen in humans supplementing with curcumin, which similarly stimulates AMPK activity.[27] In a 2019 meta-analysis published in Frontiers in Pharmacology, researchers concluded that curcumin is effective in improving metabolic health, mainly through reducing weight and leptin levels.[28] However, given the bioavailability issues inherent in curumin, how practical seeing such results is questionable.
With significantly superior bioavailability and similar action, DHZ is a more effective AMPK-activator than curcumin. This, combined with curcumin showing the ability to fight weight gain and improve insulin sensitivity in humans, leads us to believe that there is great potential for DHZ as a metabolism-boosting, blood sugar-improving ingredient.
Linoleic acid
While overall energy balance largely influences weight and fat mass, there are aspects of our diet that can be equally as integral to our metabolic health. One such factor is the ratio of omega-6 fatty acids to omega-3 fatty acids in the diet.

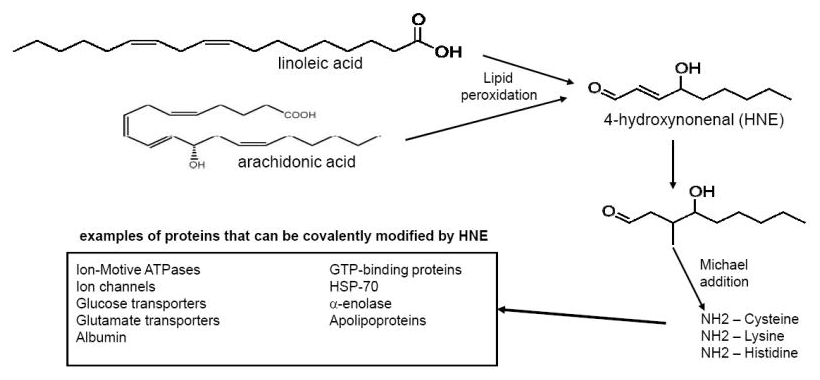
Linoleic acid itself may not be the actual problem, but its oxidation products leading to the deadly HNE quite possibly are![29] But... dehydrozingerone prevents oxidation of linoleic acid!
Polyunsaturated omega-6 fatty acids — such as those found in nuts, corn oil, and soybean oil — are somewhat common in the processed foods eaten in a standard Western diet. While their sheer presence isn't inherently a bad thing, their ubiquity leads to an overconsumption that is — research shows that consuming a high ratio of omega-6 to omega-3 fatty acids (fish, seeds, nuts) can lead to metabolic dysfunction, in addition to various diseases and conditions.[30,31] An imbalanced diet, swung too heavily in favor of omega-6, increases the risk of obesity and obesity-related issues like inflammation.[31]
The mechanism at work behind omega-6 oxidation is almost as scary as its prevalence. Linoleic acid, a major omega-6 fatty acid found in various processed fats and oils,[30] is oxidized into into 4-hydroxynonenal (HNE) via the proinflammatory omega-6 arachidonic acid and the endocannabinoid 2-arachidonoylglycerol (2-AG).[29,32,33] This chain encourages endocannabinoid hyperactivity, which has been shown to increase cravings and food intake and adiposity.[33] The oxidation of linoleic acid results in an increased desire to eat, which certainly isn't advantageous to someone looking to keep off unwanted weight.
Balancing out the ratio of omega-6s to omega-3s, thus, is an incredibly important factor to metabolic function, weight, and overall health. Eating more omega-3 fatty acids in an effort to balance total fatty acid consumption can help keep things in check, of course, but DHZ presents another avenue toward addressing omega-6 overconsumption. A 2011 study published in the European Journal of Medicinal Chemistry found that DHZ can inhibit oxidation of linoleic acid.[34] This attenuates the stimulation of appetite-inducing endocannabinoids at bay, limiting the damage that excess omega-6s can wreak on the body.

Dehydrozingerone's Effects with respect to its chemical structure.[35]
ZinjaBurn — potent DHZ extract from NNB Nutrition
In our fat burner, we're opting for ZinjaBurn from NNB Nutrition. Possessing all of the premier ingredient forumator's signature characteristics — 100% purity, lab-tested, unadulterated — this ingredient has all of the anti-inflammatory, adipose tissue-blocking capabilities that DHZ possesses.
Though it delivers additional benefits (such as radical oxidation and even mood enhancement)[36,37] ZinjaBurn is used here mainly to stimulate AMPK activity. This increases energy demands and glucose tolerance, promoting overall metabolic function. Toss in efficient fatty acid oxidation, and this NNB Nutrition ingredient packs a metabolic punch perfect for our formula.
Safety and dosing

Looking to try this potent ginger extract? NNB Nutrition's ZinjaBurn is the way to go!
DHZ can be isolated from both Zingiber officinale rhizomes (ginger roots) and Aframomum giganteum (another ginger family plant),[38,39] creating extracts that pass the 1994 DSHEA classification of a "constituent of a botanical" eligible for use. Additionally, this ingredient has received generally recognized as safe (GRAS) status (under the name of "vanillylidene acetone"),[40] and has been declared safe by the Joint FAO/WHO Expert Committee on Food Additives.[41]
For supplementation, NNB Nutrition recommends two servings per day, with between 400 to 600 milligrams per serving. Our formula consists of three 133.33 milligram servings, good for a daily 600 milligram yield. While this dose puts us on the outskirts of what's recommended, it's a careful choice — 400 milligrams of ZinjaBurn is still effective in its own right, and we had to make room for other exciting fat-burning ingredients we wanted to include!
To learn more, read our articles titled Dehydrozingerone: Ginger's Forgotten Anti-Inflammatory Weight Loss Factor and ZinjaBurn by NNB Nutrition: The Anti-Inflammatory Adipose Tissue Assassin!.
-
Caffeine Anhydrous - 400mg (133.3mg per capsule)
Caffeine anhydrous fights a two-front war in our fat burner, providing energy and a metabolic boost. Caffeine is one of the most well-researched and understood supplements in sports nutrition, showing potency in raising energy levels and improving performance in athletic training.[42]
The mitochondria are the powerhouses of our cells. But how do they work, how is our food supply damaging them so badly, and what can we do to fix the issue? Prepare to meet the Power of Mito, presented by NNB Nutrition.
In regards to weight loss, this stimulant does quite a lot. Not only does it boost mental and physical energy (both of which can help give you the energy you need to train harder and progress faster), but it also suppresses appetite and raises internal body temperature.[43,44] These effects actually come from the same action that yields energy — adenosine inhibition.[43,44]
Caffeine also downregulates phosphodiesterase, which is an enzyme that breaks down cyclic adenosine monophosphate (cAMP).[43,44] Because cAMP helps facilitate the metabolic breakdown of lipids and carbohydrates,[45] caffeine is able to prolong its activity and keep the metabolic fire burning.
We're using 400 milligrams of this energy-boosting, fat-burning stimulant here, split evenly across three servings. In our suggested supplementing protocol, that amounts to roughly 267 milligrams in the morning, 133 milligrams midday. You need to be cognizant of both your stimulant tolerance and other stimulants you decide to use in conjunction with this formula — if your tolerance isn't all that high, then taking two capsules in the morning alongside a big cup of coffee may not be the best idea! Similarly, make sure you're taking the third capsule early enough in the afternoon that it won't mess up your sleeping habits!
-
GlucoVantage (Dihydroberberine) - 200mg (66.7mg per capsule)
While ramping up the metabolism increases the body's demand for food, how the body responds to calories is just as important — this is what makes insulin sensitivity so crucial to our health and body composition. Insulin is a hormone secreted by the pancreas that moves glucose from the bloodstream and into muscle cells (for muscle repair and growth) and fat cells (for fat storage).[46-48] Having higher insulin sensitivity means that cells respond extremely well to insulin secretion, and only small amounts of the glucose-moving hormone is needed to lower blood sugar.
There's no better GDA ingredient than berberine, and there's no better form of berberine than dihydroberberine in NNB's GlucoVantage!
Insulin sensitivity largely influences the body's ability to respond to the foods we eat. A body capable of effective nutrient partitioning is one that uses calories primarily for fuel and minimizes storage. This is particularly useful when looking to shed body fat — carbohydrate-rich meals require a larger amount of insulin,[49] which in turn can inhibit the body's ability to oxidize fatty acids.[49,50] By boosting insulin sensitivity and nutrient partitioning, one allows the body to handle those carbs fast and get back to burning fat.
Dihydroberberine - superior insulin response
With high insulin sensitivity being advantageous to keeping off unwanted body fat (and insulin resistance being a marker of metabolic syndrome),[51] means by which one can improve their insulin response become attractive in a fat-burning supplement. Dihydroberberine (DHB) is one of many different nutrient-partitioning agents out there, with clinical data suggesting that it may be among the best.
To truly understand the astounding capabilities of DHB, we need to look at the data supporting berberine — DHB is simply a metabolite of berberine, acting in similar pathways while possessing a significantly important quality we'll get to soon.
Berberine can do a lot for metabolic health, but...
According to a study published in Biochimica et Biophysica Acta in 2006, researchers found that berberine stimulates the phosphorylation of AMPK,[52] the metabolic-regulating protein we discussed earlier. This heightened activity causes downstream benefits, one of which is improved insulin sensitivity.[52]
A 2012 study published in Evidence Based Complementary and Alternative Medicine advanced this finding, looking deeper into the auxiliary benefits a berberine-facilitated improved insulin response could potentially yield. They found that berberine:
How does the best glucose disposal ingredient in berberine get any better? It's known as dihydroberberine, and sold as NNB Nutrition's GlucoVantage, and if there's one anti-aging ingredient we suggest, it's this one!
-
Inhibits adiponectin and leptin expression in preadipocytes.[53] This suggests that berberine can suppress the body's initial process of fat storage. Adiponectin isn't all bad in terms of insulin response, however. Optimal balance of the three adiponectin multimers — trimer (LMW), hexamer (MMW), and high-molecular-weight (HMW) — can actually improve insulin sensitivity. In a 2011 study published in FEBS Letters, berberine was shown to promote adiponectin multimerization, raising HMW adiponectin levels while simultaneously suppressing overall adiponectin expression.[54] There is actually a duality to berberine's metabolic health-promoting effects via adiponectin activity.
-
Lowers blood glucose, hemoglobin, triglyceride levels, and cholesterol.[53] All of these are important markers of metabolic health, thus leading the researchers to suggest that berberine could be an effective treatment for metabolic syndrome.
In a 2008 study published in Metabolism, researchers tested berberine against metformin, a leading diabetic drug, in volunteers with type 2 diabetes. After three months of treatment (at equivalent doses), they found that while both groups significantly improved their blood glucose uptake capabilities, those taking berberine saw superior results.[55]
Why choose DHB over berberine? Quality and bioavailability!
All of this sounds great, of course, so the question then becomes why opt for DHB instead of berberine? Well, berberine has two main knocks against it — poor quality and poor bioavailability!
Want to get more berberine with less side effects? Dihydroberberine / GlucoVantage makes it happen!
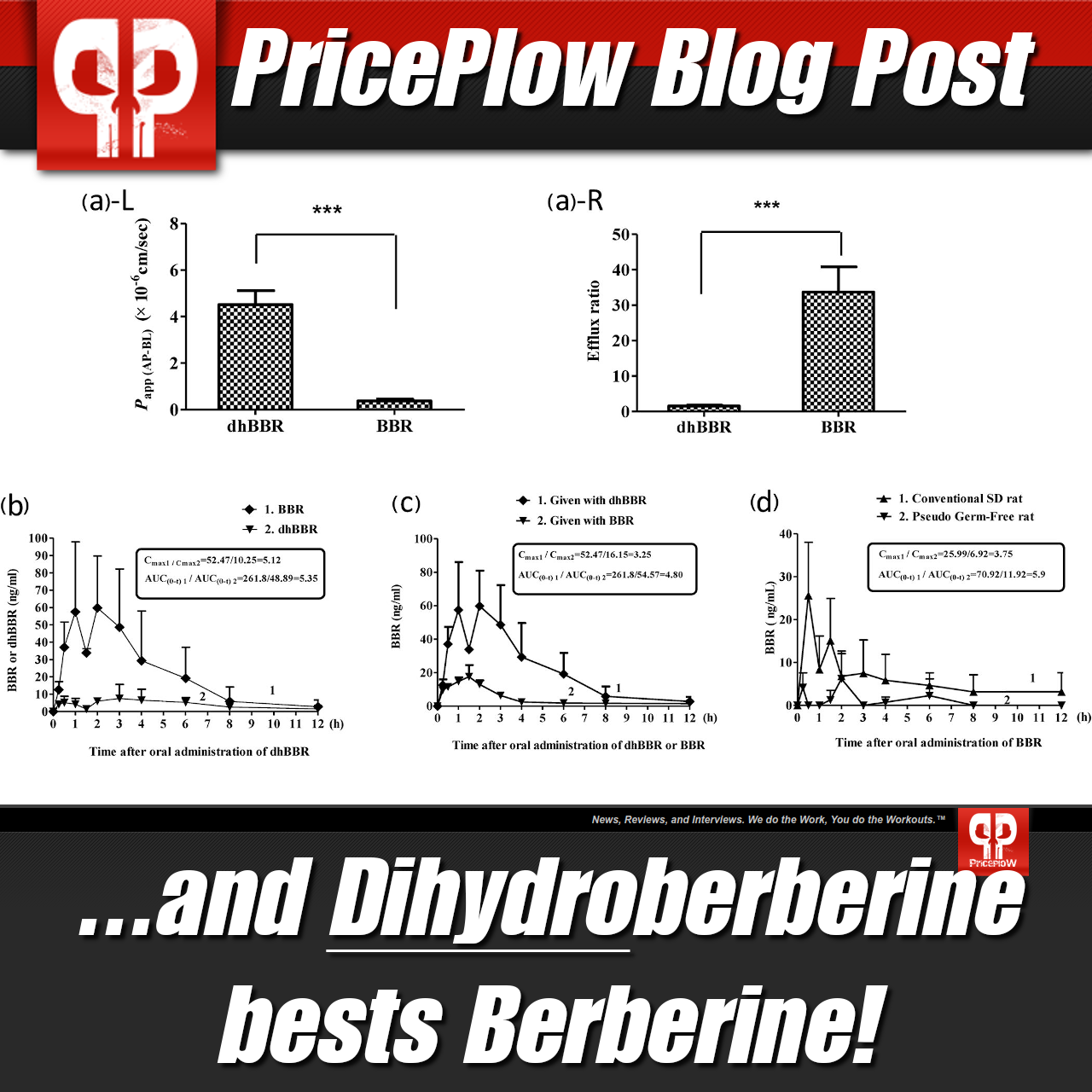
It's incredibly difficult to get real, unadulterated berberine in practice. In 2015, NNB Nutrition's chief science officer, Shawn Wells, tested 40 berberine supplements and found that 37 of them were underdosed and/or spiked. Now consider the amount needed to yield results. Many of the studies testing berberine have had to use massive doses to see significant effects, sometimes noting serious gastrointestinal discomfort alongside treatment.[55] These two circumstances make it incredibly difficult for a formulator to use berberine — it's hard to trust, hard to procure, expensive, and impractical for most supplement formulas!
DHB circumvents these issues. Its chemical structure makes it more readily absorbed into the intestine,[56] where it's then oxidized back into berberine and sent out into the blood.[56] In other words, DHB acts as a "Trojan Horse" of sorts, moving through an impasse as one thing before unveiling itself as another. A 2015 study published in Nature News compared the DHB to its parent compound in this regard, finding that DHB had an absorption rate five times higher than that of berberine.[56]
Thus, lower doses of DHB measure up against higher doses of berberine in terms of effectiveness. In a 2008 study published in Diabetes, researchers tested the two ingredients in mice — one group was given 100 milligrams per kilogram body weight of DHB daily, while the other received 560 milligrams per kilogram body weight of berberine daily — for two weeks. They found that those treated with DHB had significantly improved markers of adiposity and triglyceride accumulation compared to the berberine group, while seeing a 44% increase in insulin response.[57]
GlucoVantage — the industry’s premier DHB ingredient
In order to capitalize on the potent blood sugar-optimizing capabilities of DHB, we again turn to NNB Nutrition. GlucoVantage, part of their Vantage line of ingredients, is the industry's first commercially available DHB supplement. This non-GMO, unadulterated ingredient is derived from natural berberine and brings with it all the benefits DHB has shown in research — more insulin sensitivity, improved blood sugar, and body composition support.

GlucoVantage from NNB Nutrition is a pure form of Dihydroberberine (DHB) and is known as "The Super Berberine".
As a pure DHB ingredient, GlucoVantage delivers these effects in a more efficient manner, boasting superior bioavailability and increased potency than berberine. This makes it a perfect addition to our formula, providing strong insulin sensitivity-boosting benefits in a serving size much smaller than what we'd have to use if opting for berberine instead.
As for that serving size, NNB Nutrition recommends 100 to 200 milligrams, taken one to two times per day. In our fat burner, we're going with a total of 200 milligrams split between three capsules — that means you'd get roughly 134 milligrams in the morning and 66 milligrams midday. This keeps the blood sugar-optimizing compound at work in the body, effectively partitioning nutrients and allowing the body to not only put your daily calories to use, but to encourage a fat-burning state, too.
To learn more, read our articles titled Berberine: The Best Glucose Disposal Ingredient Just Got Better and GlucoVantage: Dihydroberberine for Superior Insulin Sensitivity.
-
-
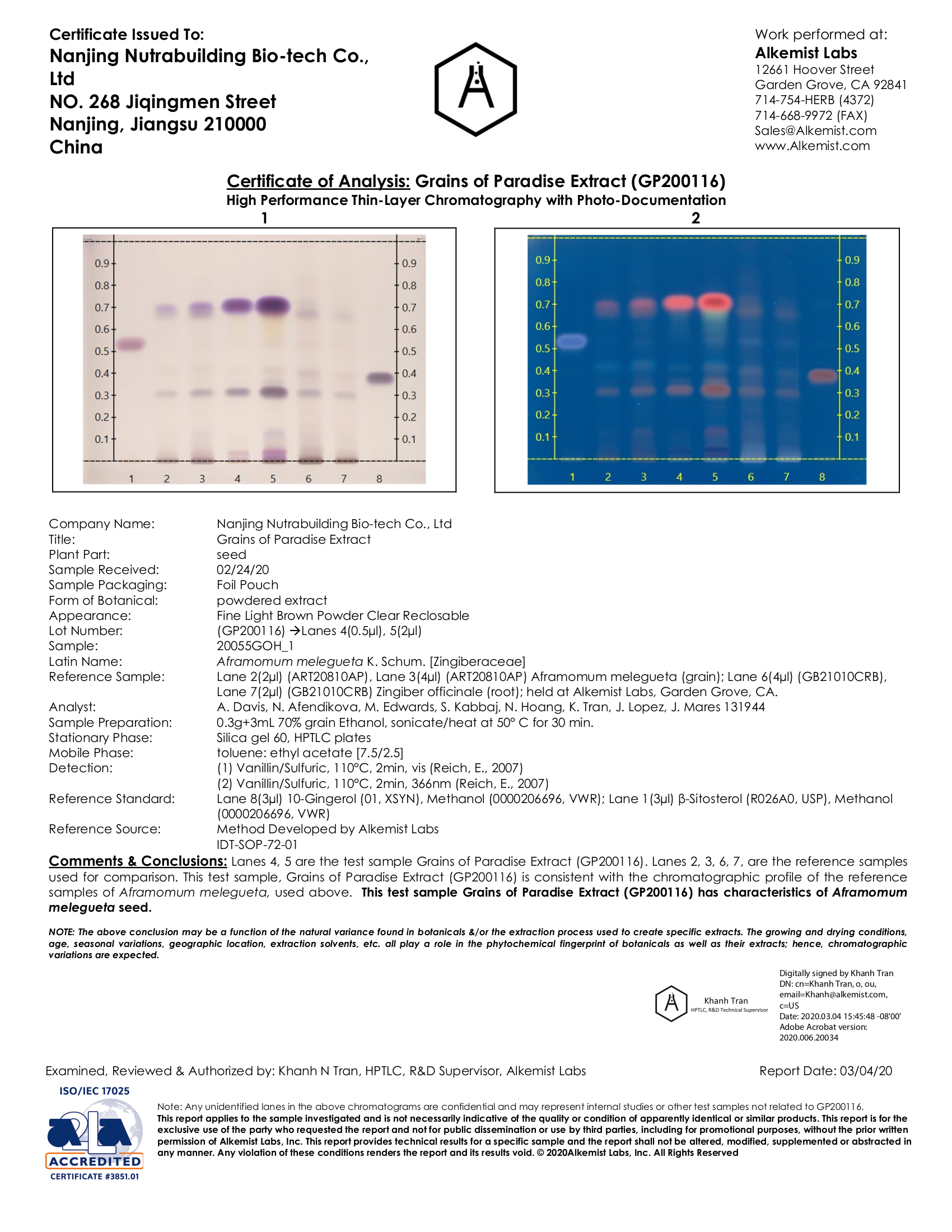
CaloriBurn GP (Grains of Paradise Seed Extract) (6-Paradol 12.5%) - 120mg (40mg per capsule)
Grains of paradise (GP), also known as Aframomum melegueta, is yet another plant native to the ginger family. This plant has a long history in folk medicine as a means to treat various ailments, with some cultures even using it as a cooking spice to add some heat to their foods.
Finally, a grains of paradise extract that actually passes lab tests for all clinical constituents!
The plant's main bioactive compounds — 6-gingerol, 6-paradol, 6-shogaol, and 6-gingerdione — aren't just the constituents that help deliver that culinary spiciness, but they also lie at the center of many of the plant's biological capabilities. Most notably, these compounds stimulate peroxisome proliferator-activated receptors (PPAR),[58] an action we covered earlier when discussing BAIBA. Activating PPAR helps turn white fat into "beige fat",[2,6] converting it into a brown fat-like type of tissue primed for energy expenditure. Furthermore, GP seems to target this energy-spending fat once it creates it, using a different mechanism from other spices like capsaicin.[59]
In a 2011 study published in Autonomic Neuroscience, researchers administered a standardized GP extract and measured discharges from nerves entering brown adipose tissue. They found that GP raised nerve activity for as long as three hours, while also increasing its temperature.[58] This action yields rather simple results — it encourages the body to burn it off in order to maintain normal internal body temperature. In other words, GP activates BAT and kickstarts the metabolism, encouraging it to burn more body fat.
BAT-activated metabolic boost in humans
In a 2014 edition of the Journal of Nutritional Science and Vitaminology, a team of researchers split 19 healthy volunteers into two groups. One group received a 30 milligram per day dose of GP, while the other received a placebo. This procedure was followed for 30 days, with the volunteers' total energy expenditure and visceral body fat being metrics of interest.

Researchers found that the group consuming the Grains of Paradise extract had a significantly greater increase in energy expenditure, due to increased BAT activity, than the group not receiving the GoP extract.[60]
They found that the group given GP saw significant increases in daily energy expenditure and decreases in visceral fat.[60] Importantly, these metrics were statistically significant both between individual volunteers and groups — GP induced these effects at the volunteer-by-volunteer level, as well as on the group level. The researchers found that the GP group ended up burning, on average, 100 calories more per day than the placebo group. That might not sound like a lot, but this additional expenditure can add up over time and really boost your fat-burning efforts.
A full-fledged extract works best, but 6-paradol plays a huge role
While each bioactive constituent in GP — 6-gingerol, 6-paradol, 6-shogaol, and 6-gingerdione — are important in their own right, research has found that 6-paradol is a key driver of much of what GP has the capability to do.
In the aforementioned 2011 Autonomic Neuroscience study, an isolated 6-paradol extract was tested alongside GP. Interestingly, they found that mice injected with 6-paradol saw similar increases in nerve activity and temperature in BAT compared to the GP group.[58] This study highlights the action of 6-paradol, unlocking the thermogenic benefits of a full-fledged GP extract.

This third-party lab result confirms that we have true grains of paradise, with at least the four peaks we also want.
With this importance understood, most GP extracts used are standardized for 6-paradol content. In the majority of the successful human-based trials, the GP used holds a 12.5% 6-paradol concentration, in addition to delivering the plant's other bioactives.
CaloriBurn - a standardized, trusted GP extract
CaloriBurn, the third ingredient in NNB Nutrition's Burn series (joining MitoBurn and ZinjaBurn), is one of the industry's highest-quality grains of paradise extract available. It's validated via multiple chromatography tests for its bioactive constituents, and is standardized to the clinically-suggested 12.5% 6-paradol content that has been so effective.
As is commonplace for NNB Nutrition ingredients, CaloriBurn is naturally-sourced and completely unadulterated. Though we always love to see such transparent, well-manufactured ingredients, these traits are even more noteworthy in this case. Unfortunately, there are "grains of paradise" available that are not real GP — some manufacturers have been accused of taking raw ginger extracts and adding 6-paradol to it. If these accusations are true, not only are these products non-compliant, but they also fall short in effectiveness.
NNB Nutrition doesn't just push the limits of ingredients in terms of innovation, but also in quality. This is evidenced by CaloriBurn, a pure grains of paradise extract that gives the industry a variant of the fat-burning spice that's both potent and trusted, with a verified content of all of its bioactive constituents.
Safety and dosage

See the lab work above: NNB Nutrition's CaloriBurn is our preferred grains of paradise extract!
GP, due in large part to its historical use as a spice, holds GRAS status that applies to its supplemental use, as well. Research also supports its safety, with studies finding virtually no side effects tied to GP supplementation.[61]
In our formula, we're using CaloriBurn to supply a total of 120 milligrams of grains of paradise — 80 milligrams in a two-capsule morning dose, 40 milligrams in a single capsule taken midday. This will help spice things up, raising internal body temperature to encourage the burning of BAT and improvements in overall caloric expenditure and body composition.
To learn more, read our articles titled Grains of Paradise: Fat Fighting Spice of the Goddesses and CaloriBurn: NNB Takes Grains of Paradise to Hot New Heights!.
-
Bitter Orange (Citrus Aurantium) [Fruit] Extract [Std. to 30% p-Synephrine] - 250mg (25mg synephrine per capsule)
Last in our fat-burning formula is synephrine.
Found in Citrus aurantium, also known as bitter orange, synephrine is a protoalkaloid that acts as a stimulant in the body. This phenylethylamine derivative is often related to other compounds with a similar structure that have been shown to have some adverse effects, but this extrapolation is unjustified — synephrine does have similar action, but it acts without bringing along those unwanted side effects (when used responsibly, of course).
Synephrine binds to ꞵ-3 adrenergic receptors in adipose tissue,[62] triggering activation of the cAMP-PKA axis and increasing the body's ability to breakdown fats.[63] Notably, this action does not influence heart rate or blood pressure — this is what separates synephrine from other structurally-similar compounds, no adverse effects on the cardiovascular system.[62] In terms of what it does do, synephrine is all about helping ignite fat-burning. Research has found that synephrine can raise basal metabolic rate and suppress appetite.[62]

CaloriBurn preserves all of the metabolism-enhancing bioactive compounds from aframomum melegueta
That being said, using this ingredient in conjunction with your weight loss-focused exercise can take things a step further. In a 2016 double-blind, placebo-controlled study published in the British Journal of Clinical Pharmacology, researchers tested the acute effects of synephrine in 18 healthy volunteers prior to exercise. They found that those who received the ingredient saw significant increases in fat oxidation rate during exercise compared to placebo, effectively elevating the fat-burning potential of their training.[64]
In a 2015 edition of the Journal of the International Society of Sports Nutrition, researchers found that both synephrine on its own and synephrine taken alongside caffeine raised fat oxidation during exercise.[65] This synergy is oftentimes capitalized upon in sports supplements, as is the case in our formula!
We're using a total of 75 milligrams of a regulated synephrine extract, standardized for 30% p-synephrine content. This ensures we're getting an effective dose of the bioactive protoalkaloid that delivers these potent fat-burning effects. This dose is split into thirds, of course, and by our protocol, you'd get 50 milligrams in the morning, 25 milligrams midday. This is enough to work in unison with everything else in the formula to keep BAT activated and fed to the metabolism for fat-burning energy.











How to Take Our Fat Burner
Another look at the label - 2 capsules will be seriously heated, and we'd take three total in an entire day, but not all at once
We don't have many stacking suggestions for our fat-burning formula, as it contains a comprehensive panel of ingredients designed for aiding weight loss. Of course, you can use it in conjunction with your normal training supplements — especially those peri-workout — so long as you're aware of the timing of stimulant consumption and your overall stimulant tolerance.
That being said, a weight loss effort is best embarked upon when the body is functioning efficiently. That means you should have a hold on your daily vitamin and mineral intake, nutrition, and stress/sleep. Adding a daily multivitamin to the mix would be a smart move, and if needed, a stress-reliever and/or sleep aid.
Otherwise, taking two capsules with your morning beverage and one more capsule midday is how we'd take this product!
PricePlow’s Spin on Fat-Burning
Dieting is tough work — it takes optimal training, nutrition, and sleep, in addition to incredible amounts of dedication and discipline to reach weight loss goals. Once these variables are all accounted for, introducing a supplement designed to help such efforts can get you to the finish line faster, perhaps with better results.

In our first Formulator's Corner installment, we built a better MCT Creamer using ingredients the rest of the market is missing - chiefly, C8Vantage MCT Powder!
We've come up with this concoction as a means to aid dieters and others looking to drop excess weight and achieve a leaner physique. While there are plenty of great fat burners out there already, we wanted to create a small label that leveraged multiple high-quality ingredients (alongside some stimulants, of course) to raise fat-burning potential. This formula takes aim at the mechanisms that drive fat-burning ability and body composition — brown adipose tissue activation, metabolic boosting, insulin sensitivity optimization — to provide a comprehensive diet aid.
Of course, calling on four of the premium, innovative ingredients from NNB Nutrition allowed us to draw up this formula. With these ingredients in tow, alongside the synergistic pair of caffeine and synephrine, we feel pretty confident about the ability of this formula to kickstart the metabolism and help shed unwanted body fat.
Sign up for our future NNB Nutrition news, and after that, you can see a couple single-ingredient products that use some of the ingredients discussed above:
Subscribe to PricePlow's Newsletter and Alerts on These Topics
Alpha Lion Gains Candy GlucoVantage – Deals and Price Drop Alerts
Get Price Alerts
No spam, no scams.
Disclosure: PricePlow relies on pricing from stores with which we have a business relationship. We work hard to keep pricing current, but you may find a better offer.
Posts are sponsored in part by the retailers and/or brands listed on this page.
Alpha Lion Gains Candy CaloriBurn – Deals and Price Drop Alerts
Get Price Alerts
No spam, no scams.
Disclosure: PricePlow relies on pricing from stores with which we have a business relationship. We work hard to keep pricing current, but you may find a better offer.
Posts are sponsored in part by the retailers and/or brands listed on this page.

Comments and Discussion (Powered by the PricePlow Forum)