UNBOUND has officially burst onto the supplement scene, with "exciting" being the best word to describe the launch. Prior to the big unveil, everyone in sports nutrition wondered who was drumming up all this intrigue. The inauguration — complete with a drum containing all six flagship supplements — validated the hype. It was revealed that CEO Mark Glazier and his team at NutraBio were the ones behind Unbound, and it all made sense. Their experience, knowledge base, and drive to innovate are woven through the fabric of this new endeavor.

UNLOCK your fat loss with this potent fat burning drink!
Now that we've covered both Unbound's pre-workouts — the heavy-hitting, euphoric UNLOAD and the powerful, pump-inducing non-stimulant formula UNBENT — it's time to discuss one of the most exciting, powerful formulas in the entire product line.
It is time to UNLOCK your fat loss
Just like its pre workout brethren, UNLOCK also happens to be one of the most comprehensive, potent supplements in its category -- this time fat burning drinks. This fat-incinerating formula is absolutely loaded — 11 active ingredients, six of which are trademarked products from leading formulators — and ready to help unlock the next level of fat loss for those deep in the throes of a diet.
Before we get to this stacked label, make sure you're subscribed to PricePlow for the latest news and deal alerts so that you can stay up-to-date on all things Unbound. Also, head over to our social feeds on Instagram, YouTube, and Facebook, for daily content on new products, supplement reviews, and some informative Q&As!
Unbound Unlock – Deals and Price Drop Alerts
Get Price Alerts
No spam, no scams.
Disclosure: PricePlow relies on pricing from stores with which we have a business relationship. We work hard to keep pricing current, but you may find a better offer.
Posts are sponsored in part by the retailers and/or brands listed on this page.
This area is reserved for Team PricePlow's upcoming videos.
Subscribe to our channel and sign up for notifications so you catch it when it goes live!
![]()
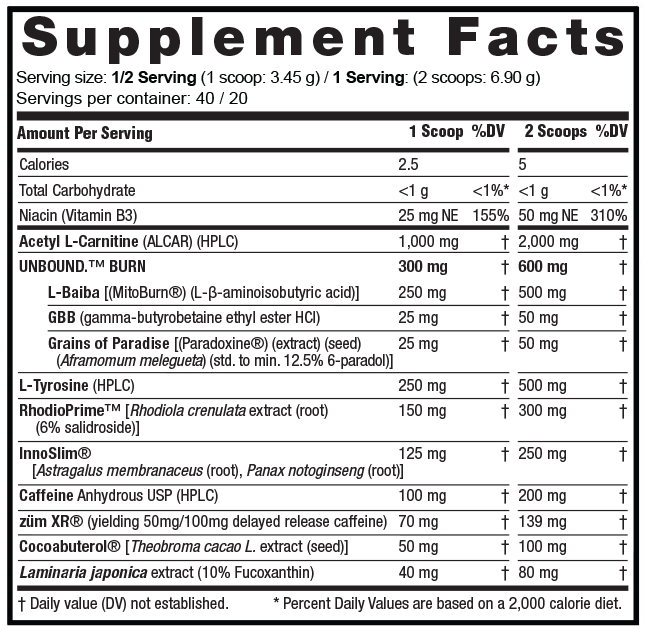
Ingredients
UNLOCK lists two suggested serving sizes: 1 scoop (3.45 grams) and 2 scoops (6.90 grams). Because they identify 2-scoops a full serving, we're going to cover the label using that dose.

UNLOCK the door to fat loss with Unbound's incredible metabolic-enhancing fat burning drink
Here's what you get get in each 2-scoop serving:
-
Acetyl L-Carnitine (ALCAR) (HPLC) - 2,000mg
Unbound formulates their products using a combination of the more staunch sports supplement ingredients and those that lean toward innovation. The first ingredient in UNLOCK, L-carnitine, firmly sits in the former group — it's one of the most heavily used ingredients in weight-loss and fat-burning products.
How carnitine works

A closer look at the incredible Unbound UNLOCK Ingredients label. Click to zoom in
Carnitine frequents these types of products primarily due to its biological action in the body. It's a conditionally essential amino acid, which means that the body can manufacture it on its own when needed. It's commonly found in various proteins, such as chicken, dairy, and importantly, red meat. Whether consumed through food or synthesized endogenously, the body absolutely needs it. Stored mainly in skeletal muscle, carnitine is used to metabolize fatty acids.[1,2]
The endogenous carnitine pathway takes place mainly in the liver and kidney and begins with
L-methionine and L-lysine, two essential amino acids.[3] Four enzymatic reactions reduce these amino acids first to trimethyllysine (a methylated form of lysine) before yielding circulating carnitine that can be used by the body.[3,4] This process is facilitated by multiple key nutrients, such as ascorbic acid (vitamin C) and niacin (vitamin B3),[3] the latter of which is included in UNLOCK. That being said, this is more of a complement than anything else — exogenous carnitine, or carnitine consumed through the diet, is available right away.

After reading a new review based upon 100 citations, we are finding fewer and fewer reasons not to take ~2g L-Carnitine each day
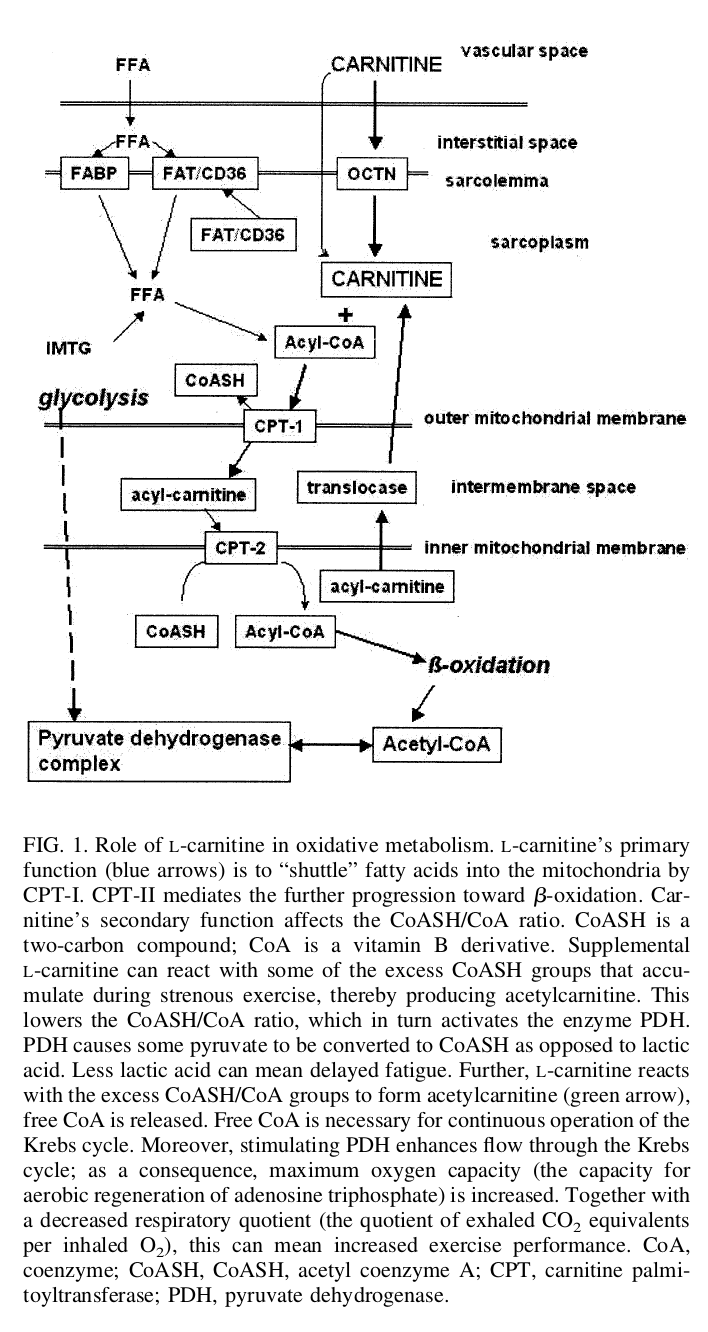
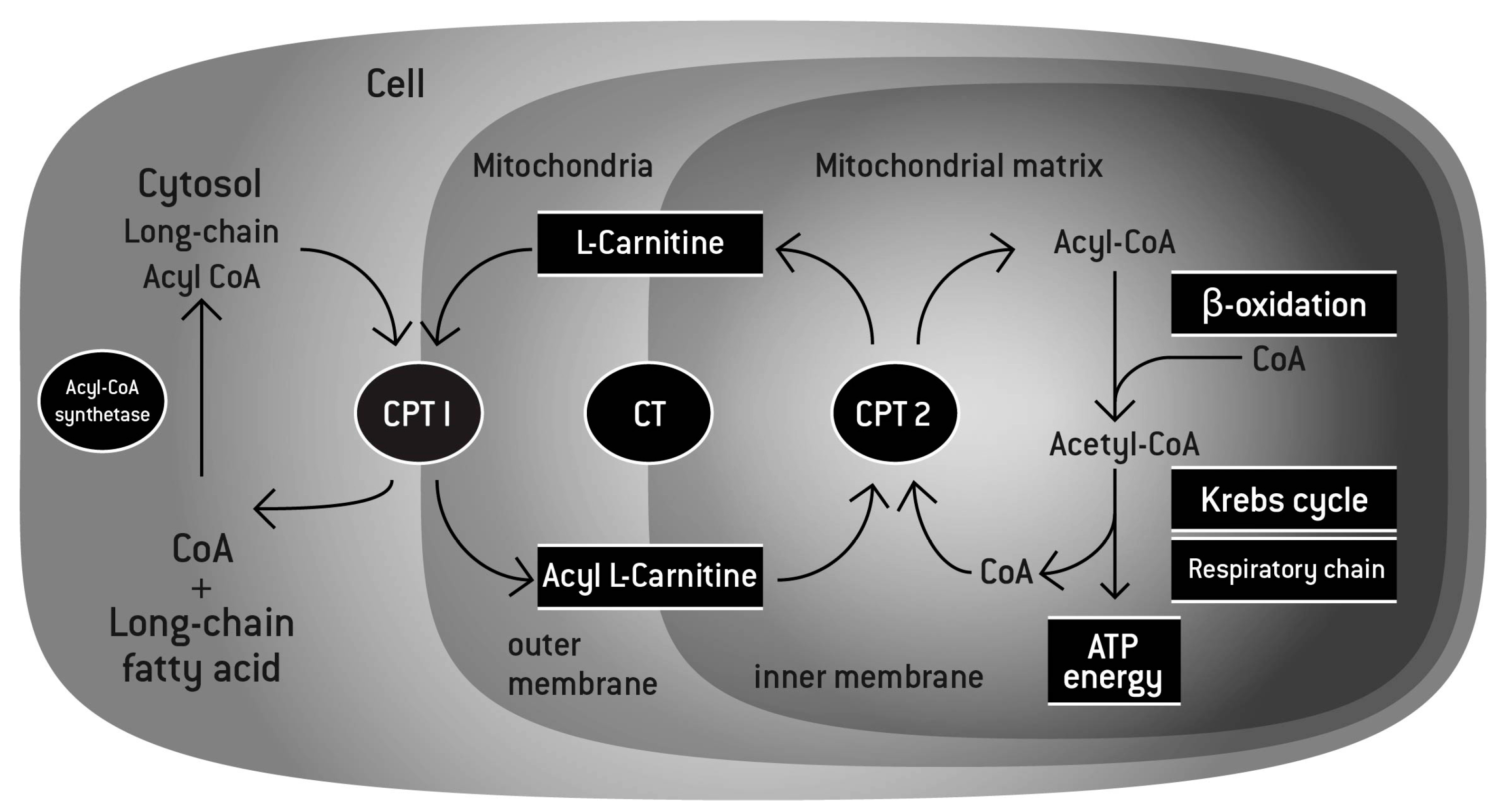
Carnitine consumed through food gets absorbed in the intestines by the organic cation transporter 2 (OCTN2), a key sodium-dependent transporter in the digestive system.[5] Dispersed directly into the bloodstream, carnitine begins to go to work — operating through carnitine palmitoyl transferases (CPT1 and CPT2),[3,5] it binds to long-chain fatty acids and moves across the mitochondrial membrane — notably, the mitochondria are the parts of the cell that create energy.[3] From there, carnitine encourages the beta-oxidation of the fatty acid it's bonded to, effectively producing acetyl-coenzyme A (Acetyl-CoA), a central figure in the TCA cycle and the production of adenosine triphosphate (ATP), in the process.[3] In essence, carnitine moves fatty acids into the cells so that they can be burned for energy, making this amino acid a key regulator of metabolic functioning and energy homeostasis.
Boosts metabolic health
Having such an effect on metabolic functioning, it stands to reason that carnitine would influence metabolic health just as much. In a study published in 2009, individuals consuming 2 grams of carnitine per day for six months saw significant improvements in insulin sensitivity and glucose response.[6] These effects have been shown to be even more pronounced in individuals with glucose intolerance and insulin secretion issues.[7]
Notably, carnitine is capable of supporting efficient metabolic firing when used in conjunction with exercise. In a 2011 study, researchers gathered 14 healthy individuals who engaged in recreational athletics and trained between three and five times per week. They were split into two groups— one received two 80-gram carbohydrate drinks a day, while the other was given two 80-gram carbohydrate drinks containing 2 grams of carnitine in each. Various metrics were tested over three visits.

L-carnitine's role in oxidative metabolism
The researchers found that after 24 weeks, the carnitine group had significantly increased skeletal muscle content by 30% compared to the placebo group.[8] They also put these higher pools of carnitine to use — in an exercise performance test (30 minutes of "all-out" exercise plus 30 minutes of moderate exercise), those that had consumed carnitine performed 35% more total work than the control group. Such results were supported by lower levels of muscle lactate and spared muscle glycogen content.[8] Carnitine effectively oxidized fatty acids for more usable energy, allowing the athletes to maintain performance and burn more energy in the process.
A 2018 review supports the benefits of carnitine supplementation in the metabolic functioning of athletes, allowing them to perform at higher levels. In assessing the results of more than 30 studies, these researchers found conclusive evidence of carnitine's ability to enhance exercise capacity and endurance,[9] despite seeing mixed results in terms of changes in VO2 max, lactate buildup, and muscle growth.[9] This review concludes that carnitine can help improve exercise performance and recovery.[9]
But what about fat mass?
Unfortunately, the research is a bit less definitive in terms of fat loss. In a 2016 review of nine randomized, controlled clinical trials, carnitine showed the ability to aid weight loss and lower body mass index (BMI) compared to control groups.[10] However, these results run contradictory to a 2000 study in which moderately obese volunteers who took a carnitine supplement and participated in an aerobic training program saw no significant difference in weight or fat mass after eight weeks.[11]
What this tells us is, as always, you have to do the work. The biochemical workings are true and reliable — carnitine facilitates the beta-oxidation of fatty acids — but this doesn't necessarily mean it simply burns fat. Now, leveraging a carnitine supplement alongside an intense training program and a quality diet? That might do it — not only will the amino acid support the metabolism, but it will assist in maintaining the training intensity necessary to reduce fat mass and improve body composition.
Nootropic benefits

L-carnitine function, as described above
Supplemental carnitine can come in a few forms, though L-carnitine and acetyl-L-carnitine (ALCAR) are most common. ALCAR is simply carnitine bonded to an acetyl group, and while this may seem trivial, it actually opens another door...one that leads to nootropic action.
ALCAR is capable of crossing the blood-brain barrier, inducing a cognitive effect that, according to multiple studies, can improve mental capacity,[12,13] promote neurotransmitter production,[14] and provide neuroprotective and anti-depressive benefits.[15]
Enhanced levels of focus and clarity, no matter how small, can go a long way — particularly for those who are dieting and combating the "diet fog" that can accompany eating below energy maintenance levels.
UNLOCK kicks things off with an ingredient that supports a fat-loss effort in two ways — ALCAR not only promotes fat-burning at the cellular level but also improves cognition, two things that are extremely useful when on a diet. At a big 2 grams of a high-performance liquid chromatography (HPLC), lab-verified variant of the amino acid, this formula packs more than enough to leverage both of these actions.
-
UNBOUND BURN - 600mg
The Unbound BURN matrix is a three-pronged fat-burning powerhouse that utilizes multiple patented ingredients to really ignite the metabolism. While these inclusions are often found in fat burners and are quite potent, it's not all that often we see them used together. But because Unbound looks to smash complacency, they've decided to use effective doses of all three.
-
L-BAIBA (MitoBurn) (L-β-aminoisobutyric Acid) - 500mg
MitoBurn L-BAIBA has flipped the fat burner niche on its head!
L-ꞵ-aminoisobutyric acid, also known as BAIBA, is an amino acid that exists as a signaling molecule in the body. A natural byproduct of muscle contraction — and generated through the breakdown of the amino acids valine and thymine — this myokine communicates that contraction to the rest of the body.[16] Specifically, BAIBA is released from skeletal muscle in response to an exercise stimulus, triggering multiple bodily processes that take place when we exercise.[16]
Signaling PPAR
Many of the mechanisms initiated by BAIBA pertain to the metabolism and begin with the peroxisome proliferator-activated receptor-γ coactivator-1⍺ (PGC-1⍺). This transcriptional protein is activated in response to exercise and is responsible for facilitating operations like mitochondrial biogenesis, enhanced insulin response, fatty acid oxidation, and thermogenesis.[17] However, PGC-1⍺ is restricted to myocytes and cannot travel across the body — it relies on key messengers, such as irisin and BAIBA, to transmit its activity elsewhere.
Following a rise in PGC-1⍺ activity, BAIBA prompts peroxisome proliferator-activated receptor-γ (PPAR-γ) and peroxisome proliferator-activated receptor-⍺ (PPAR-⍺), two nuclear receptor proteins that regulate fatty acid oxidation and adipocyte differentiation.[16,18,19] Furthermore, BAIBA also signals uncoupling protein-1 (UCP-1) and cell death-inducing DFFA-like effector A (CIDEA), two genes expressed in brown adipocytes.[19]
Brown fat cells form brown adipose tissue (BAT), which is the type of fat that the body uses for energy.[20] Through a combination of PPAR and UCP-1 upregulation, BAT is fed into the respiratory chain and readily oxidized by the metabolism.[20] Contrast this type of fat tissue to white adipose tissue (WAT), which is the visceral fat that not only regulates leptin levels (the hormone that controls appetite), but is difficult to burn off.[20]

Beta-aminoisobutyric acid (BAIBA), induces beneficial effects on lipid homeostasis in mice.[21]
This comparison highlights why BAIBA is such an exciting ingredient. A study published in 2014 found that BAIBA activates UCP-1 and CIDEA via PPAR-⍺ activation in white adipocytes.[16] In essence, BAIBA facilitates the "browning" of WAT into "beige" fat — this conversion allows for the altered tissue to be oxidized more efficiently and used for energy. Rising PPAR-⍺ activity has effects elsewhere, too, as it also increases fatty acid release in the liver to further encourage fat-burning.[22]
Encourages ketone synthesis
Speaking of fatty acids and the liver — beta-hydroxybutyrate (BHB) is a ketone manufactured in the liver in response to fatty acid oxidation[23]. The production of BHB serves as an important intermediate in the relationship between hepatic fat oxidation and metabolic function.[24]

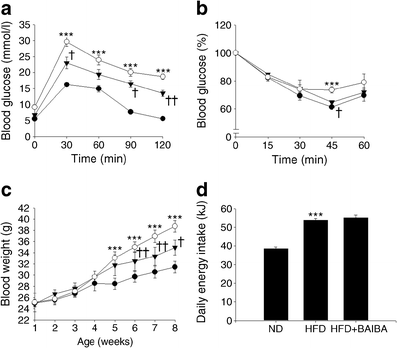
Despite eating the most calories, the high-fat diet mice that also received BAIBA had far less weight gain and better blood glucose scores than their non-BAIBA counterparts.[25]
In a 2003 placebo-controlled trial, researchers fed a BAIBA-infused diet to mice and assessed their metabolic health. Two different doses of BAIBA — either 10 milligrams per kilogram of body weight or 100 milligrams per kilogram of body weight — were used, with the mice consuming their assigned diet on a daily basis for two weeks. They found that the 100-milligram group saw increased beta-oxidation of fatty acids, facilitated by a 64% increase in total ketone bodies.[26]
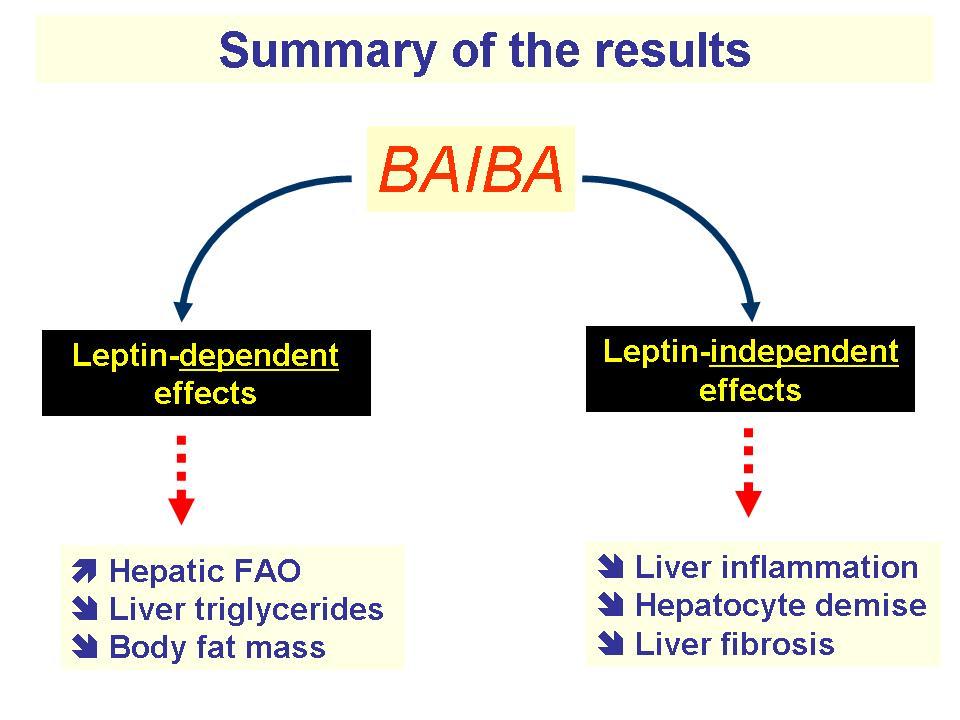
Another study, conducted in 2004, supported these findings. Mice treated with BAIBA saw increased BHB levels.[27] However, this study took things a step further, inspecting the effects such an increase has on body fat. They found that despite zero changes in diet, other than BAIBA supplementation, these mice experienced significant decreases in fat mass compared to baseline.[27]
BAIBA-modulated BHB production helps empty hepatic fat stores, effectively allowing the body to utilize other fatty acids for oxidation. As such, BAIBA encourages fat-burning by clearing out the liver and tapping into fat stores elsewhere.
Hormonal effects — leptin and insulin
In addition to influencing protein receptors and ketone bodies, BAIBA impacts two hormones that are integral to metabolic health.
Leptin is a hormone stored in WAT that controls the sensation of satiety and energy balance. Research shows that in individuals with higher leptin levels — a side effect of added fat mass — leptin sensitivity decreases.[28] This means it's more difficult for individuals with more fat mass to listen to satiety cues and lose weight, even if they're dieting.

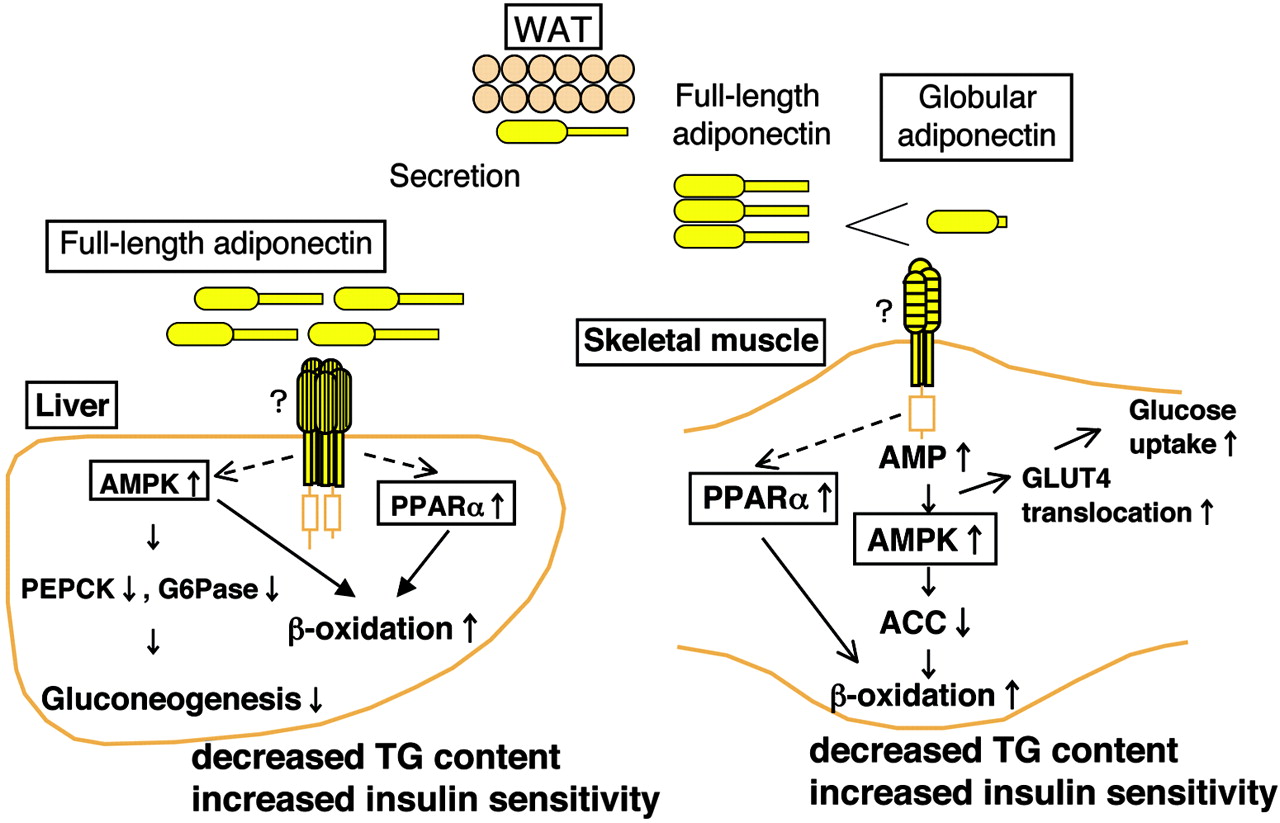
Adiponectin can activate AMPK and PPARα in the liver and skeletal muscle. We recently explored PPARα in our articles on BAIBA and MitoBurn.
A study from 2012 found that mice treated with a daily dose of BAIBA (either 100 or 500 milligrams per kilogren of body weight) for four months avoided body fat gain when fed a high-calorie diet.[21] The underlying effects of BAIBA explained why — not only did the researchers see higher plasma BHB levels, but they also observed enhanced leptin sensitivity.[27
Insulin, a hormone secreted by the pancreas, largely controls glucose response by transporting sugars from the blood into myocytes, adipocytes, and the liver. Insulin resistance — a condition in which the body struggles to respond to insulin and fails to efficiently use glucose in the blood for energy — is a key indicator of metabolic dysfunction, as well as various other diseases and conditions.
In the previous study in which BAIBA improved leptin levels, the researchers also saw the amino acid improve glucose tolerance.[21] Two other studies — published in 2015 and 2016 — reached similar results. BAIBA supplementation effectively reduced glucose intolerance and improved insulin sensitivity in mice.[25,29]
MitoBurn — NNB Nutrition's innovative L-BAIBA ingredient
BAIBA is a relatively new ingredient in sports supplements, with research highlighting its potential but few capitalizing on it. Whenever there's a chance to innovate and break conventions in the industry, one particular ingredient formulator circles that opportunity more than others — NNB Nutrition.

NNB Nutrition has finally brought us a trusted and tested form of L-BAIBA, which we call an "exercise signal" that kickstarts incredible metabolic processes! It's known as MitoBurn and it helps kick-start the 'exercise program'!
NNB Nutrition is leading the charge in bringing BAIBA to market with their groundbreaking MitoBurn. This ingredient utilizes L-BAIBA, the bioactive form of BAIBA that research suggests is responsible for signaling metabolic processes (D-BAIBA, the other form, doesn't have much support in this regard).[30,31]
Of course, MitoBurn comes with all the characteristics of an NNB Nutrition product — it's unadulterated, lab-tested, and standardized — in addition to the effectiveness we've come to expect from their ingredients. Taking all of this into consideration, MitoBurn presents one of the best ways to introduce BAIBA into your fat-burning regimen.
If you're interested in reading more about L-BAIBA, make sure you check out our previous coverage on both the ingredient itself and MitoBurn (when you're finished reading about UNLOCK, of course!):
- BAIBA: New Weight Loss Ingredient Generates Exercise in a Pill?!
- MitoBurn: β-Aminoisobutyric Acid (L-BAIBA) from NNB Nutrition
NNB Nutrition recommends taking MitoBurn at a dose between 250 to 500 milligrams, once or twice daily. UNLOCK sits on the high-end of that range, providing an ample 500 milligrams of this potent BAIBA variant. This inclusion will work to expedite fatty acid oxidation by "browning" WAT for readily available energy, facilitating fat loss in the process.
-
GBB (Gamma-Butyrobetaine Ethyl Ester HCl) - 50mg
Gamma-butyrobetaine (γ-butyrobetaine, or GBB) is a naturally occurring metabolite of L-carnitine. An intermediate in the enzymatic breakdown of trimethyllysine into carnitine (which we touched on earlier), GBB is reduced by gamma-butyrobetaine dioxygenase (BBD) into carnitine.[4] Notably, this conversion takes place specifically in the liver, kidneys, and brain — the only places where BBD is active in the body.[32]

GBB Supplements add to the "carnitine pool" by increasing the reagent -- GBB -- which goes on to make more carnitine if your body needs it!
GBB metabolizes directly into carnitine, representing a key relationship between the two — the body tries to maintain an appropriate balance between both molecules. Through a variety of enzymatic processes, the body metabolizes GBB into carnitine (and vice versa) to maintain homeostasis between the two. In essence, GBB facilitates carnitine production, effectively boosting whole-body carnitine levels to further leverage the benefits we discussed earlier in relation to this amino acid.
Raises carnitine levels
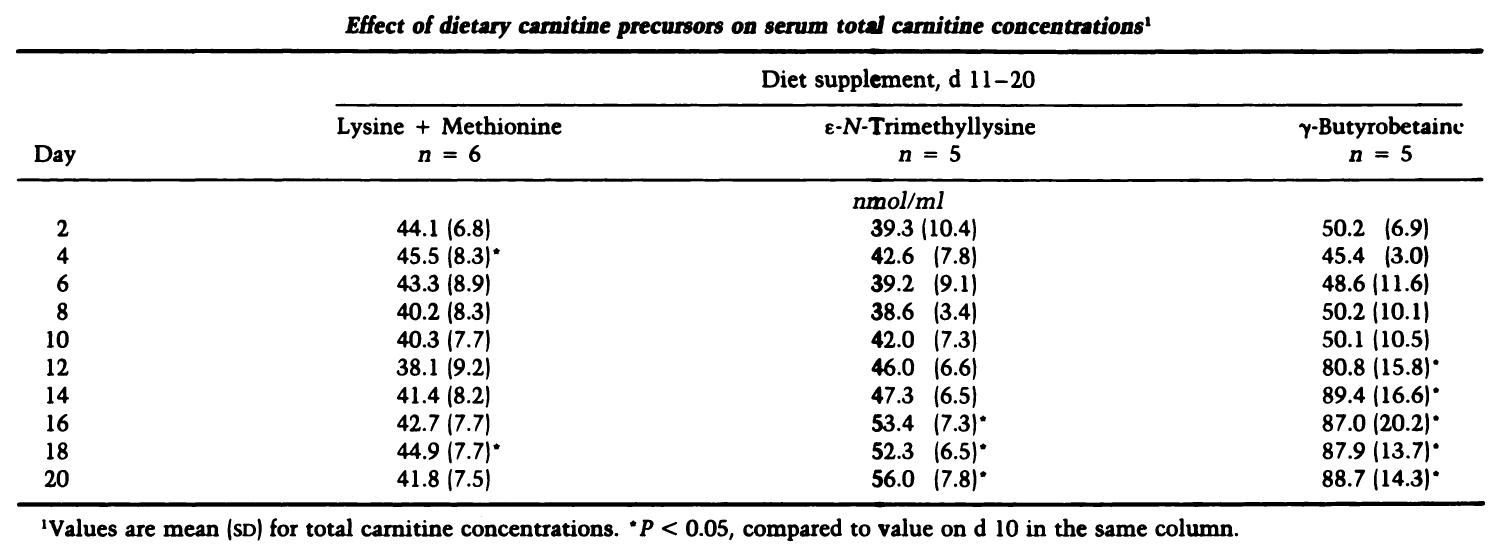
Given its place in carnitine metabolism, it should come as no surprise that GBB increases carnitine in the body — the degree to which it does so, however, is impressive. Two clinical trials using individuals on a low-carnitine diet — one used infants while the other gathered adults — introduced a GBB supplement to see how it affected plasma carnitine. In both cases, the results were astounding — plasma carnitine levels doubled, in addition to a three-fold increase in carnitine excretion.[33,34]
It's important to note that the individuals in these studies were consuming low levels of carnitine through their diet. People who consume large amounts of high-carnitine foods (such as red meat), or supplement with carnitine (such as with a product like UNLOCK) likely wouldn't see such a pronounced increase. That being said, it could still act as a complement, further elevating carnitine levels and leveraging the associated benefits.

The pathway image above is more detailed, but the point is that your body will do its best to maintain hemeostasis and can go in both directions
Get sweaty!
In addition to clinical data, GBB seems to have a heat-producing effect. Though this action is reliant on anecdotal sources, it does make sense — GBB increases carnitine levels, which in turn elevates fatty acid oxidation and thus, increases thermogenesis. Generating more internal heat leads to more sweating, a natural way for the body to cool itself.

An early study on carnitine's precursors showed that GBB (to the right) is the best of the bunch at increasing plasma carnitine levels.[34]
As such, GBB is often used in supplements to turn up the heat, adding a bit of sweat to a formula in addition to promoting carnitine production.
Coming in the form of GBB ethyl ester HCl — GBB that's capable of being ingested and metabolized by the body — this formula uses 50 milligrams. According to clinical data, this is an appropriate dose, especially considering everything else UNLOCK delivers.
-
Grains of Paradise (Paradoxine) (Extract) (Seed) (Aframomum Melegueta) (Std. to Min. 12.5% 6-Paradol) - 50mg
Now it's time to spice things up. Grains of paradise (GP), also known as Aframomum melegueta, is a spice native to the ginger family that has a history around the globe, both as a culinary ingredient and an herbal remedy.

They're not just for cooking anymore! In this article, we discuss how the proper extract from these seeds can be used for weight loss and body recomposition!
Plants in the ginger family are touted for their health-promoting properties, which are driven by powerful bioactive compounds that vary across the genus. GP generally contains four phenylpropanoid-derived compounds, but this extract focuses primarily on 6-paradol, which is what is used in the studies and gives the plant some of its spicy taste as well as some of its biological action. This compound primarily stimulates peroxisome proliferator-activated receptors (PPAR),[35] which as we covered earlier, converts white adipose tissue into a "beige fat" that serves as a more accessible source of fuel for the metabolism.[18,19]
This effect turns up the heat, increasing thermogenesis and raising body temperature. In a 2011 trial, mice fed a standardized GP extract had the activity of the sympathetic nerves entering brown adipose tissue (BAT) measured. The researchers saw that GP elevated nerve activity for as long as three hours,[35] while simultaneously increasing body temperature.[35] Effectively stimulating BAT, GP revs the metabolism and heats-up production.
Increases caloric expenditure and improves body composition
In 2014, researchers gathered 19 human volunteers and split them into two groups — one received 30 milligrams of GP daily, while the other was given a placebo. This double-blind procedure was carried out for 30 days, with metrics pertaining to energy expenditure and body fat of particular interest to the research team.

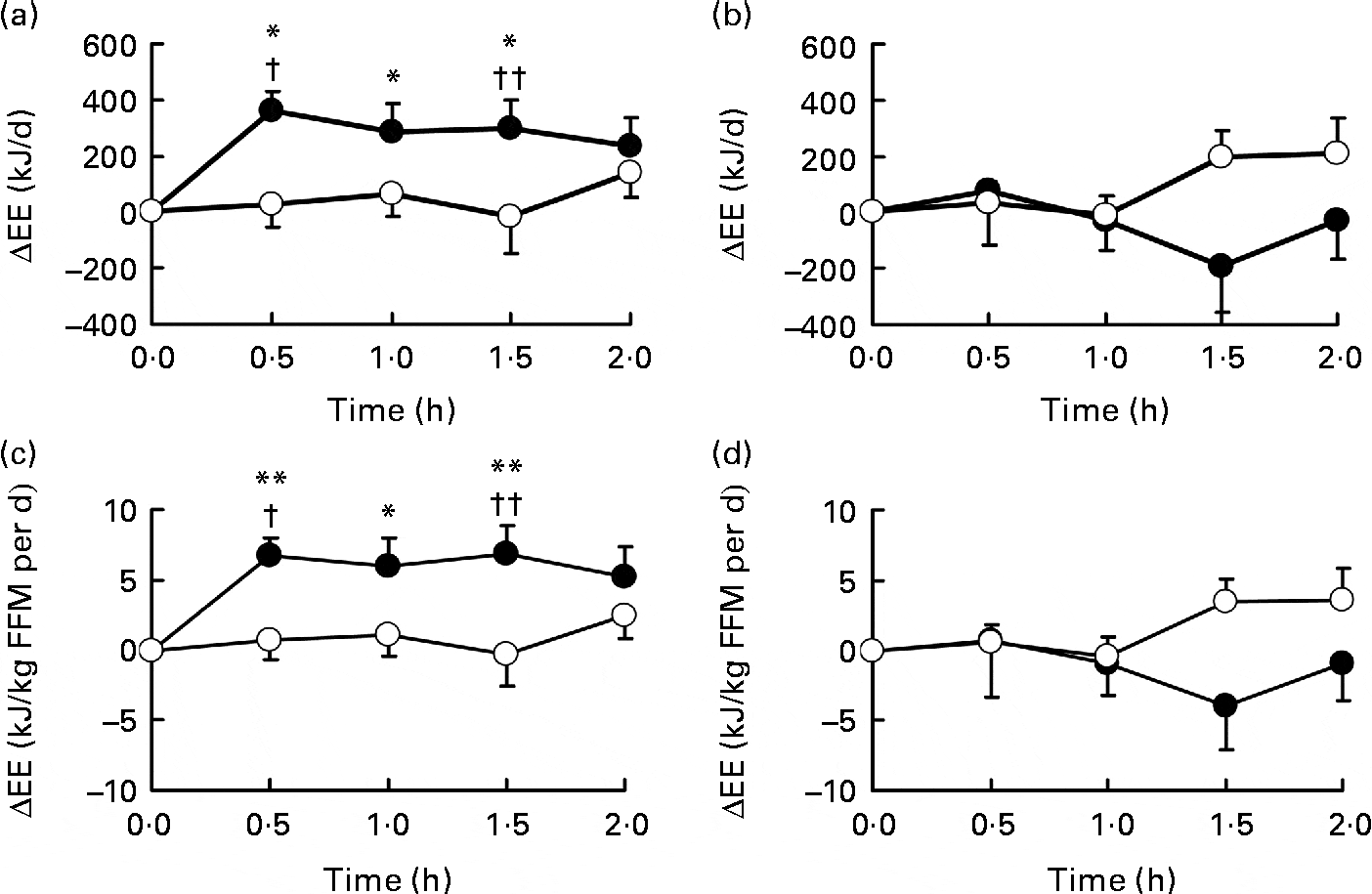
Energy expenditure change (ΔEE) after oral ingestion of Grains of Paradise.[36] This spice helps melt away those stubborn pounds!
At the conclusion of the study, they found that the GP group saw both increases in daily calorie expenditure and decreases in visceral body fat.[36] These results were both significantly better when compared to baseline measurements and the results of the placebo group at the study's conclusion, as well. Those assigned to GP treatment burned, on average, 100 calories per day more than the placebo group[36] — a notable increase that many dieters would surely appreciate.
GP standardized for 6-paradol
With research showing that 6-paradol, taken by itself, can induce the same BAT-stimulating effects as a full-fledged GP extract.[35] It's important to have a comprehensive GP ingredient, but with 6-paradol acting as the key to unlocking the ingredient's full potential, any quality GP extract is standardized for this specific gingerol. Paradoxine is one of these quality extracts, having been used for years in sports supplements. Standardized for a clinically-backed 12.5% 6-paradol, this ingredient delivers all the potential GP brings to the table.
UNLOCK utilizes a hot 50 milligrams of this heat-generating spice, which has yet to show any adverse effects when taken by healthy individuals.[37] At one time, this dose was considered to be on the higher end, and still is in powdered formulas that must be flavored. It now represents a moderate and effective hit, capable of heating things up.
-
-
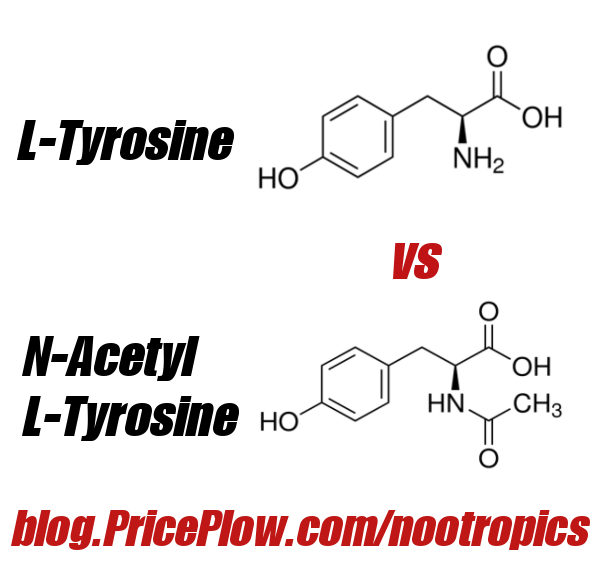
L-Tyrosine (HPLC) - 500mg
Tyrosine is a non-essential amino acid found naturally in high-protein foods such as dairy, poultry, and fish. It's grown into a popular ingredient in sports supplements for its ability to improve cognition and facilitate metabolic health, two capabilities that are incredibly relevant in the case of UNLOCK.

Long story short from our analysis -- For the best effects from tyrosine, choose the regular L-Tyrosine version!
Tyrosine is a precursor to dopamine, norepinephrine, and epinephrine, a group of key neurotransmitters known as catecholamines. These chemicals facilitate numerous functions in the brain by enabling cellular signaling. Individually, they have some important effects:
- Dopamine regulates focus, mental sharpness, motor function, and motivation.[38] Playing a vital role in modulating mood, dopamine is especially useful in maintaining a calm, positive demeanor.[39] Dieting can be tough — so more dopamine can help reduce the perceived burden.
- Norepinephrine and epinephrine act separately but are often discussed together due to their synergy in regulating the fight-or-flight response.[40,41] Stimulating this system raises heart rate and lipolysis, in addition to suppressing appetite upon acute activation.[42]
This slight involvement with appetite leads us to a larger metabolic effect — tyrosine, alongside the mineral iodine, promotes proper thyroid function. These two compounds facilitate the synthesis of triiodothyronine (T3) and thyroxine (T4), two hormones secreted by the thyroid that control various metabolic processes.[43,44] The goal here isn't necessarily more thyroid hormone production but rather homeostasis. Proper thyroid function — which balanced hormone levels promote — is correlated to healthy body weight and energy expenditure.[45] Tyrosine helps keep this gland balanced, encouraging a well-functioning metabolism in the process.
With the ability to improve metabolic function while fighting the potential mood and motivation issues that accompany a difficult weight-loss endeavor, a high quality, HPLC-verified tyrosine is a great inclusion in this formula. At 500 milligrams, considered the lower end of the range suggested by research, it's still a clinical dose. We've seen higher doses of tyrosine (such as in UNLOAD, which packs a heavy 3 grams of the nootropic), but we're content with what we have here.
-
RhodioPrime [Rhodiola Crenulata Extract (Root) (6% Salidroside)] - 300mg
Speaking of UNLOAD, that formula and UNLOCK share this phenomenal ingredient in common. Just as UNLOAD is the first pre-workout to use RhodioPrime 6X from NNB Nutrition, UNLOCK is the first fat burner to use it, too!
RhodioPrime 6X from NNB Nutrition is a trusted source, and high in salidroside!
Its purpose here is similar, as well. Rhodiola is an adaptogen with a long history in herbal medicine, with various cultures and practices using it for mood improvement, vitality, and longevity, among other things.[46] The plant's bioactive compounds — salidroside and rosavin — are responsible for this wide array of anecdotal applications, as science has supported what was previously passed on through word-of-mouth.
With salidroside being tabbed as the driving constituent behind Rhodiola, research has found that the herb can increase long-term potentiation (LTP) in the hippocampus,[47] promote cellular function,[48] stimulate catecholamine activity,[49-52] and improve amine uptake in the brain.[49-52] These all improve cognitive function and focus, two effects that, as we've said, can be highly valuable to dieters.
In a double-blind, placebo-controlled clinical study published in 2000, researchers tested just how impactful Rhodiola could be in this regard. They treated college students studying for finals with a daily dose of 100 milligrams of the herb for 20 days. Compared to placebo, the Rhodiola students showed significant improvements in cognitive function, mental sharpness, and general well-being.[53] These findings were corroborated by a similar study from 2004 in which stressed-out college students experienced similar "brain gains" following a one-time dose of Rhodiola.[54] Both test groups were dealing with stressful situations, yet managed to improve their mental clarity. Dieting introduces both mental and physical adversity, which gives these studies some relevance here.
What makes RhodioPrime different?
Rhodiola: The Adaptogen Powered by Salidroside. In this article, we dive deep into rhodiola, and take a different approach to the adaptogenic herb!
Not only is Rhodiola an under-used ingredient in fat burners, but this particular variation is even more unique. RhodioPrime 6X is a standardized Rhodiola crenulata extract formulated by NNB Nutrition to contain at least 6% salidroside. This characteristic places it far above most other Rhodiola extracts when comparing concentrations of the integral bioactive. An effective, HPLC-verified, high-quality Rhodiola extract, RhodioPrime 6X further pushes this product's cognitive effects to new heights.
UNLOAD delivers 300 milligrams of this focus-enhancing agent, which is the dose suggested by NNB Nutrition and far more than we see in weight loss supplements that care to include it. This one's going to feel good.
-
InnoSlim [Astragalus Membranaceus (Root), Panax Notoginseng (Root)] - 250mg
InnoSlim, formulated by NuLiv Science, is a patented blend of Astragalus membranaceus and Panax notoginseng and has been clinically shown to improve metabolic function. This ingredient modulates the metabolism by affecting multiple proteins, enzymes, and cofactors that play a significant role in facilitating its operation.
Reduces SGLT1 activity...
More than ever, we understand metabolism and its critical importance. Besides exercise and cleaning up your diet from processed foods, how else can we accelerate our path back to metabolic wellness? Nuliv Science's InnoSlim is one such accelerant.
In a 2007 in vitro study, researchers treated human Caco-2 cells with InnoSlim. The team was mainly focused on the ingredient's effect on glucose absorption — Caco-2 cells (which live in the digestive tract) uptake glucose via SGLT1, a sodium-dependent transporter that moves sugars from the digestive system into the blood.[55] They found that InnoSlim reduced Caco-2 glucose absorption by 42% compared to placebo, inhibiting SGLT1 activity by 21% in the process.[56] This means that InnoSlim reduces the release of sugar into the bloodstream, which can significantly benefit insulin sensitivity, blood glucose levels, and overall metabolic health.
...while increasing GLUT4 activity!
Though inhibiting SGLT1 in the gut can limit the rate at which sugars enter the blood, stimulating GLUT4 can limit how long they remain there. GLUT4 is the body's main glucose transporter, operating mainly in the blood where it moves sugars to skeletal muscle and adipose tissue.[57]
In 2008, NuLiv funded a study similar to the one previously discussed. This time, however, instead of Caco-2 cells, researchers administered InnoSlim to 3T3-L1 adipocytes (fat cells) and C2C12 myotubes (muscle cells). They found that InnoSlim increased glucose uptake by 142% and 43% in fat and muscle cells, respectively,[58] modulated by a spike in GLUT4 activity.[58] This action not only keeps blood glucose levels in check but also facilitates recovery by shuttling glucose to the cells that need it.
Affects multiple metabolism-associated genes and enzymes
In addition to its effects on these two glucose transporters — which have a significant influence on metabolic health — InnoSlim has an impact elsewhere, too. Other in vitro studies have found that InnoSlim:
- Suppresses glycogenesis and lipogenesis in liver cells.[59] This can help defend against liver dysfunction, a marker of metabolic syndrome.[59]
- Increases AMP-activated protein kinase (AMPK), acetyl coenzyme A carboxylase phosphorylation (ACC-P), and adiponectin.[59] All three of these components are vital to effective fatty acid oxidation, glucose uptake, and ultimately, metabolic efficiency.[59-62] Adiponectin is also a key modulator of appetite and body weight, with its activity limited in the presence of higher levels of body fat.[63-65]
Clinical backing

Since InnoSlim is non-stimulatory, the doors are opened to a multitude of incredible weight loss applications.
Seeing just how potent InnoSlim was in over 18 in vitro and animal studies, NuLiv Science funded a study using human volunteers. Two groups were split in a double-blind manner — one was given InnoSlim (at an undisclosed dosage) while the other was given a placebo. All subjects were told to maintain their lives as normal — the same eating habits, activities, and so on — while taking their assigned treatment.
The research team saw that those who received InnoSlim displayed a 30% increase in adiponectin expression compared to placebo,[66] which accordingly raised AMPK, GLUT4, ACC-P, and overall caloric expenditure.[66] This finding (plus reduce inflammation, improved glucose uptake, and healthier blood triglyceride levels) explained a more noteworthy finding — the InnoSlim group saw, on average, significant reductions in body weight and body fat percentage compared to placebo.[66]
Just one of many powerful NuLiv Science ingredients
We've covered NuLiv Science for years at this point, and they continue to impress by expanding their ingredient profile. One of the industry's leading ingredient formulators, NuLiv Science has developed an array of ingredients that offer benefits ranging from blood glucose support, nutrient absorption, cellular regeneration, and even improved skin health!
What's more — they tend to use proprietary extracts that leverage either (or both) astragalosides or ginsenosides, the two main bioactive constituents found in astragalus and notoginseng, respectively. They also leverage other herbs with strong biological activity, like Rosa roxburghii.
What are the differences in these NuLiv Science ingredients that use similar plants? We dive into the Astragalosides and Ginsenosides inside, and break down the different constituents of four of NuLiv Science's major sports nutrition ingredients
If you're interested in learning more about InnoSlim and the other ingredients NuLiv Science offers, check out the following pages:
- InnoSlim: Potent Stimulant-Free Fat Burning Ingredient by NuLiv Science
- Astragalosides and Ginsenosides: Differentiating NuLiv Science's Ingredients
Based on their data, NuLiv Science recommends that an effective InnoSlim dose sits at 250 milligrams. That's precisely what we see here, which means Unbound effectively delivers enough to leverage the blood sugar-improving, metabolism-supporting effects of this impressive ingredient. NutraBio's UpSorb notably takes advantage of their AstraGin ingredient, and would be great to stack here as well.
-
Caffeine Anhydrous USP (HPLC) - 200mg (of 300mg total caffeine)
Caffeine anhydrous is one of the more ubiquitous ingredients in sports supplements, oftentimes used to increase energy and improve athletic performance.[67] Though that applies here — and is particularly beneficial for those taking UNLOCK prior to training — caffeine actually has a direct impact on the metabolism as well, making it a great addition to a fat-burning formula.
Prepare yourself for a wild ride - Unbound's UNLOAD pre workout has several next-gen ingredients, and is the first to include RhodioPrime 6X! This also has a far higher dose of caffeine!
In addition to increasing energy and focus, which can help you push intensity in the gym, caffeine has been shown to suppress appetite and increase internal thermal temperature.[68,69] The stimulant accomplishes this by inhibiting adenosine and suppressing phosphodiesterase — the first action is central to caffeine's existence as a stimulant, while the latter prolongs cyclic adenosine monophosphate (cAMP) activity, which allows the messenger to promote metabolic processing.[70]
What does USP mean?
You may notice the "USP" abbreviation next to this ingredient and how it's not something you typically see attached to a sports supplement. The United States Pharmacopeia (USP) is a standardization that verifies the quality and purity of ingredients as set by the United States Pharmacopeial Convention. While this notation typically falls to pharmaceuticals, awarding an ingredient a "USP" label simply validates that the ingredient is of the highest-quality, most effective of its ilk. In this case, caffeine anhydrous USP means that the caffeine ingredient used here has been reviewed for potency and purity — something we can't say for the large majority of caffeine ingredients in sports supplements!
Caffeine — which, as caffeine anhydrous, is fast-hitting — contributes to almost everything this label aims to do. The stimulant increases energy, improves cognition, and facilitates fat-burning. At 200 milligrams, it's enough for an immediate jolt but is a bit below what someone looking for a hardcore fat burner might be longing for. Fear not — caffeine anhydrous only makes up two-thirds of UNLOCK's total caffeine dose!
-
zümXR - 139mg (yielding 100mg of Delayed Release Caffeine out of 300mg total caffeine)
That other one-third comes from zümXR delayed-release caffeine, a caffeine variant that has been designed to hit later, after ingestion. Its synergy with caffeine anhydrous is wonderful — anhydrous provides the energy to get you going, while zümXR kicks in just as the initial uptick begins to wane. This can help reduce the risk of a caffeine crash and prolong all of the stimulant's cognitive and fat-burning effects.
This dose of zümXR adds 100 milligrams of caffeine to the allotted total, bumping UNLOCK's overall caffeine content to 300 milligrams. This total dose is on par with many of the higher-stim products in sports supplements. While that may make it familiar for many, as always, be sure you're aware of your stimulant tolerance prior to taking UNLOCK.
-
Cocoabuterol [Theobroma Cacao L. Extract (Seed)] - 100mg
#BeUnequaled
Theobromine, a compound derived from Theobroma cacao seeds, is an extremely popular inclusion in fat-burning products. This methylxanthine is typically found alongside caffeine in nature, such as in cacao beans and green coffee beans.[71]
Theobromine is structurally similar to caffeine — both are considered methylxanthines and, thus, stimulants — and also blocks adenosine receptors, giving this compound a psychoactive effect. Interestingly, research suggests that it also regulates gene expression and decreases cellular oxidative stress, two actions that are independent of adenosine receptor blocking.[72]`
Primarily, however, theobromine is here to work in tandem with caffeine in increasing energy and fat-burning potential. Studies show that theobromine can enhance energy and cognitive function, induce lipolysis, and suppress appetite.[72,73]
Cocoabuterol is a theobromine ingredient that's standardized for at least 50% cocoa alkaloids, which are the main bioactive constituents in theobromine. UNLOCK leverages 100 milligrams of it here, a standard dose that amplifies much of what caffeine does.
-
Laminaria Japonica Extract (10% Fucoxanthin) - 80mg
Fucoxanthin is an intriguing compound found in seaweed — it actually gives seaweed a brownish color.[74] Upon ingestion, this carotenoid is deacetylated into fucoxanthinal, which is then readily absorbed via fatty acids in processes similar to that of fat-soluble vitamins.[75] Once available to the body, fucoxanthin affects multiple mechanisms, some of which we've mentioned previously. Fucoxanthin has been shown to:

UNLOCK it.
- Stimulate AMPK phosphorylation[76]
- Increase GLUT4 activity[77]
- Improve blood triglyceride levels[78]
- Increase circulating adiponectin[79]
Additionally, a study from 2009 found that fucoxanthin has a significant impact on white adipose tissue (WAT). Obese mice fed a high-fat diet were treated with fucoxanthin for five weeks, with researchers assessing various metabolically associated metrics. They found that, specifically in WAT, fucoxanthin increased mRNA expression of ꞵ3-adrenergic receptors and uncoupling protein-1 (UCP-1).[77] As mentioned when discussing BAIBA, UCP-1 activity facilitates the conversion of WAT into "beige" fat, which is more readily burned by the body. Sure enough, these two actions suppressed body weight and WAT gain in the test mice, and speak to additional research seeing enhanced fatty acid oxidation and thermal energy production.[80]
A clinical study published in 2010 provides some insight on how this ingredient may work in humans. One hundred and fifty one obese volunteers — 113 of whom had non-alcoholic fatty liver disease (NAFLD), while the remaining 38 had normal liver fat (NLF) — were given an ingredient containing 2.4 milligrams of fucoxanthin daily in a double-blind, placebo-controlled procedure. After 16 weeks, the researchers found that those assigned treatment saw significant reductions in body weight, body fat, liver fat, and blood triglycerides.[81] Notably, these results were seen in both groups — individuals with and without NAFLD saw these effects, though to varying degrees.[81]
UNLOCK rounds out its formula with 80 milligrams of Laminaria, which, standardized for 10% fucoxanthin, delivers a strong 8 milligrams of the fat-burning carotenoid. This is a pretty large dose compared to what we've seen elsewhere, and with research yet to show adverse effects,[82] we're very excited about its inclusion here.














Available Flavors
We've seen fat burners use various implements — powders, pills, and even topicals — as a delivery method. While they each have their place, some prefer adding a tasty drink to their supplement regimen, as opposed to a few capsules.
Unbound taps into their NutraBio roots — NutraBio is no stranger to making delicious supplements — in making UNLOCK powdered fat burners. They've done so with a bit of twist, too, as UNLOCK comes in a few unique flavors we've rarely seen. This makes us pretty excited to give them a try — if anyone can pull it off, it's difficult to bet against the formulators over at NutraBio!

Go Beyond Pump
Here are the flavors UNLOCK is currently offered in:
Unbound recommends mixing one serving (2 scoops) with 12 to 16 fluid ounces. If you're going to go with only 1 scoop — which may not be a bad move as you find out where your tolerance lies — adjust the amount of water accordingly!
Stack with UNBENT
Taking it pre workout? Then get the pump of your life with Unbound UNBENT, a stimulant-free pump powerhouse that hits tons of pathways to get you yoked up during and after your workout!
UNLOCK New Fat-Burning Potential
Unbound has branded themselves as a supplement company for those who are relentless in the pursuit of their goals. Their products are designed to aid in that pursuit and amplify both the experience and results. Crafted with a level of intellect, resources, and experience that's difficult to find in this industry, Unbound's initial six products certainly have the potential to help individuals #BeUnequaled.
A simple logo, but not simple supplements. Unbound is here.
This notion rings true in UNLOCK, the brand's incredibly strong, clinically-dosed, and effective fat-burning powerhouse. Utilizing 11 ingredients — including six patented ingredients from some of the industry's top formulators — UNLOCK packs an impressive amount of fat-burning potential into each serving. It won't do the work for you, of course — balanced nutrition, an effective training regimen, and other lifestyle factors will always be central to any weight-loss goal — but the market Unbound is looking at already knows that.
The only way you can reap the benefits is by putting in the necessary work. UNLOCK is for those who are already dialing in all they need to in order to get lean and healthy — these people may just be able to UNLOCK another level in their weight-loss journey, maximizing their hard work in the process.
Unbound Unlock – Deals and Price Drop Alerts
Get Price Alerts
No spam, no scams.
Disclosure: PricePlow relies on pricing from stores with which we have a business relationship. We work hard to keep pricing current, but you may find a better offer.
Posts are sponsored in part by the retailers and/or brands listed on this page.

Comments and Discussion (Powered by the PricePlow Forum)